

7)Действия на растения загрязнения среды.
Устойчивость растений к тяжелым металлам
Загрязнение среды тяжелыми металлами происходит в результате сжигания топлива, деятельности горнодобывающей промышленности, сбрасывания сточных вод и внесения в почву удобрений. Тяжелые металлы поступают в растение преимущественно через корневую систему из почвы, в меньшей степени – через листья. Скорость поглощения растением металлов зависит от рН почвенного раствора, содержания органических веществ в почве и концентрации других ионов.
Основная часть высших растений повреждается избыточным содержанием тяжелых металлов. Однако многие растения способны накапливать в основном в надземных органах большие количества тяжелых металлов, многократно превышающие их концентрации в почве. Эти растения так и называются растениями-аккумуляторами, которые в процессе эволюции, произрастая на почвах геохимических аномалий, сформировали конститутивные механизмы устойчивости к тяжелым металлам, что позволяет им аккумулировать токсичные элементы в метаболически инертных органах и органеллах или включать их в хелаты и тем самым переводить в физиологически безопасные формы. Подобные виды растений начинают активно использовать для разработки технологий биологической очистки, загрязненных территорий.
Помимо растений-аккумуляторов существуют растения-индикаторы и растения-исключатели. У растений-индикаторов содержание металла в клетках соответствует его содержанию в почве. В побегах растений-исключателей поддерживается низкая концентрация металлов, несмотря на высокую концентрацию в окружающей среде. В этом случае барьерную функцию выполняет корень.
Влияние тяжелых металлов на физиологические процессы. В основе токсического действия тяжелых металлов лежит их способность связываться с серосодержащими веществами и образовывать с ними прочные соединения. Одним из наиболее распространенных отрицательных эффектов тяжелых металлов является их взаимодействие с SH-группами белков, что приводит к инактивации ферментов и изменению других биологических свойств макомолекул и сопровождается нарушением клеточного метаболизма и физиологических процессов. В настоящее время известны более 100 ферментов, инактивирующихся тяжелыми металлами.
Наиболее общие проявления действия тяжелых металлов на растения – это ингибирование фотосинтеза, нарушение транспорта ассимилятов и минерального питания, изменение водного и гормонального статусов организма и торможение роста.
Тяжелые металлы ингибируют фотосинтез, нарушая ультраструктуру хлоропластов, тормозя синтез фотосинтетических ферментов и снижая количество хлорофилла, пластохинона и каротиноидов, вызывая дефицит СО2 из-за закрывания устьиц. Типичным результатом действия кадмия и ряда других тяжелых металлов является уменьшение содержания хлорофилла, причем концентрация хлорофилла b снижается сильнее, чем хлорофилла а. Это действие кадмия является следствием как торможения синтеза хлорофилла, так и его деградации. В присутствии таких тяжелых металлов, как Сu, Pb, Cd снижается активность ключевых ферментов фотосинтеза РуБФ-карбоксилазы и ФЕП-карбоксилазы. Кроме того, ингибируется активность карбоангидразы при действии повышенных концентраций кадмия. Тяжелые металлы влияют и на световую фазу фотосинтеза, нарушая транспорт электронов, преимущественно связанный с фотосистемой II, что обусловлено изменением структуры тилакоидных мембран, нарушением синтеза пластохинона и снижением активности ферредоксин-НАДФ+-оксидоредуктазы.
Действие тяжелых металлов на дыхание изучено слабо. Тем не менее установлено, что кадмий снижает поглощение кислорода корнями и изолированными клетками табака. Он ингибирует транспорт электронов и протонов в митохондриях, что может приводить к нарушению работы электронно-транспортной цепи. Кадмий ингибирует активность ключевых ферментов гликолиза и пентозофосфатного окислительного пути.
Под действием тяжелых металлов нарушается водный статус растения. Замечено, что многие растения в промышленных районах характеризуются меньшей оводненностью тканей и пониженной интенсивностью транспирации, что нарушает тепловой режим листа. Изменение водного статуса растения является следствием многих причин: понижения эффективности осморегуляции, уменьшения эластичности клеточных стенок, нарушения водопоглотительной способности корня. Водопоглотительная способность корня падает вследствие ингибирования формирования новых боковых корней и корневых волосков, замедления линейного роста корня, снижения контакта корневой системы с почвой, торможения транспорта ассимилятов из побегов в корневую систему. Кроме этого, ускоряется отмирание кончика корня, возрастает лигнификация и суберинизация клеток, увеличивается содержание АБК, вызывающей закрывание устьиц.
Тяжелые металлы в большинстве случаев ингибируют поглощение клетками корня как катионов, так и анионов. Торможение поглощения макро- и микроэлементов может быть обусловлено конкуренцией с тяжелыми металлами за переносчики. Другой причиной нарушения ионного гомеостаза в клетках является отток ионов (например, калия) из корней вследствие изменения под действием тяжелых металлов активности мембранных ферментов и повреждения мембран. У разных растений реакция может быть крайне неодинаковой. Так, аккумуляция меди снижалась в присутствии кадмия в корнях райграса, кукурузы, капусты и клевера, но увеличивалась в корнях риса и не изменялась в корнях тыквы и огурца.
Наиболее чувствителен к действию тяжелых металлов рост. Причем рост корня более чувствителен, чем рост побега. Кадмий и свинец сильнее ингибируют рост главного корня проростка, чем образование боковых корней, в результате корневая система приобретает компактную форму. Наиболее устойчиво к тяжелым металлам прорастание семян, что обусловлено низкой проницаемостью для них семенной кожуры. Ингибирование роста тяжелыми металлами является следствием снижения скорости как деления, так и растяжения клеток. В основе этого явления могут быть уменьшение оводненности тканей, удлинение митотического цикла, нарушение эластичности клеточных стенок и формирования микротрубочек.
Таким образом, практически все физиологические процессы в растении подвержены негативному действию тяжелых металлов. Тем не менее растения способны расти и на загрязненных тяжелыми металлами территориях, используя специальные адаптивные механизмы.
Адаптация растений к тяжелым металлам. Растения выработали целый ряд приспособительных механизмов, защищающих клеточный метаболизм от присутствующих в окружающей среде тяжелых металлов. Эти механизмы включают: связывание тяжелых металлов клеточной стенкой и выделяемыми клеткой веществами (эксудатами); снижение поступления в клетку тяжелых металлов и выброс их из цитоплазмы в апопласт; хелатирование в цитоплазме пептидами и белками; репарацию поврежденных белков и компартментацию металлов в вакуоли с помощью переносчиков тонопласта. Рассмотрим некоторые из этих механизмов подробнее (рис.).
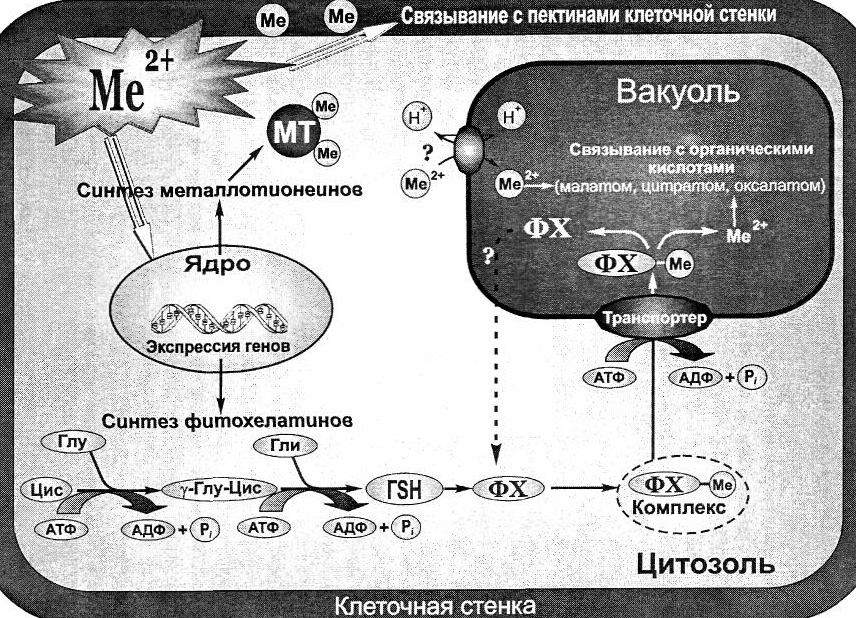
Рис. Механизмы устойчивости растений к тяжелым металлам
Клеточная стенка и корневые эксудаты. Связывание тяжелых металлов стенками клеток корня – первый рубеж «обороны» от присутствующих в почвенном растворе металлов. Цель этого механизма – снизить проникновение тяжелых металлов в протопласт. Подобный механизм функционирует, например, у толерантной к тяжелым металлам смолевки обыкновенной, которая аккумулирует их в клеточных стенках эпидермы за счет связывания с белками ли силикатами.
Выделяемая клетками и покрывающая поверхность корня слизь ограничивает проникновение тяжелых металлов в клетки, т.е. выполняет барьерную функцию. Тяжелые металлы связываются с карбоксильными группами уроновых кислот слизи.
Иногда выделяемые в почву эксудаты корней могут содержать гистидин, цитрат и другие хелаторы тяжелых металлов. Подобный механизм используется при детоксикации такого легкого металла, как алюминий. Интересно, что в данном случае у растений, например, гречихи в корнях образуется щавелевая кислота, которая не выбрасывается наружу, а поступает в листья, где алюминий аккумулируется в виде нетоксичного оксалата алюминия.
Плазмалемма играет важную роль в поддержании низкой концентрации тяжелых металлов не только за счет предотвращения или снижения интенсивности их поступления в клетку, но и за счет активного выброса их наружу. В бактериях наиболее эффективной защитой от избытка тяжелых металлов является их активный транспорт из цитоплазмы через плазмалемму. В растениях механизмы транспорта тяжелых металлов в клетку и наружу очень слабо изучены.Белки теплового шока (БТШ) не только выполняют функцию молекулярных шаперонов, но могут участвовать и в защите макромолекул и в репарации поврежденных при стрессе белков. Как уже известно, синтез белков теплового шока вызывается, прежде всего, высокой температурой, но отдельные их компоненты могут синтезироваться и в ответ на действие других стрессоров, в частности тяжелых металлов. Речь идет о группе низкомолекулярных белков (молекулярная масса 16-20 кДа) и высокомолекулярного белка БТШ-70. В культуре клеток томата БТШ-70 синтезируется не только при тепловом шоке, но и под действием кадмия. Этот белок локализован в ядре, цитоплазме и плазмалемме. Было высказано предположение, что БТШ-70 участвует в защите плазмалеммы от повреждения кадмием, так как кратковременная тепловая обработка клеток перед действием кадмия повышала устойчивость мембран к этому металлу.
Фитохелатины и другие хелаторы тяжелых металлов. Хелаты – вещества, образующие с металлом комплексную соль, в которой металл закреплен по всем валентностям и находится внутри молекулы, поэтому его возможности вступать в реакцию резко снижаются. Хелатирование металлов (образование хелатов) в цитозоле является очень важным механизмом детоксикации тяжелых металлов.
Лигандами (от лат. ligare – связывать), т.е. веществами, образующими с металлом хелат, могут служить аминокислоты, органические кислоты и два класса пептидов: фитохелатины и металлотионеины. Наиболее обстоятельно исследованы в растениях фитохелатины.
Фитохелатины (ФХ) – это пептиды, которые синтезируются в ответ на обработку растения тяжелыми металлами, активно связывают металлы и имеют структуру. Фитохелатины синтезируются не на матрице, а с помощью фермента фитохелатинсинтазы (ФХ-синтазы), использующей глутатион в качестве субстрата.
Не все тяжелые металлы одинаково эффективно индуцируют синтез фитохелатинов, и не во всех случаях имеются доказательства их защитного действия. Еще предстоит выяснить участие фитохелатинов в развитии устойчивости растений к цинку, никелю, меди и некоторым другим тяжелым металлам.
Металлотионеины (МТ) – это низкомолекулярные полипептиды, содержащие большое количество цистеина и активно связывающие металлы. Название этих соединений указывает на присутствие серы в их молекулах (греч. theion – сера). Металлотионеины были исследованы у животных и грибов. Способность растительных металлотионеинов связывать тяжелые металлы и тем самым их обезвреживать еще предстоит доказать.
В отличие от фитохелатинов металлотионеины кодируются генами и синтезируются на рибосомах обычным матричным способом. Обнаружены два класса генов, которые кодируют у арабидопсиса четыре типа металлотионеинов. Проростки арабидопсиса отвечали на присутствие меди интенсивной аккумуляцией мРНК одного из типов металлотиотеинов, но слабо реагировали на кадмий и цинк. Кроме того, установлена способность генов металлотионеинов арабидопсиса повышать устойчивость дрожжевых клеток к меди. У десяти экотипов арабидопсиса обнаружена корреляция между интенсивностью синтеза МТ2 и чувствительностью к меди.
Потенциально органические кислоты и аминокислоты (лимонная, яблочная, гистидин) являются лигандами для связывания тяжелых металлов и могут участвовать в повышении устойчивости растений. Однако прямых доказательств подобной роли этих соединений в растениях пока нет.
Компартментация тяжелых металлов в вакуолях. Выброс ионов через плазмалемму наружу или их транспорт в вакуоль – это два пути уменьшения избыточных, токсичных концентраций металлов в цитозоле. Выше уже говорилось, что аккумумуляция комплекса «кадмий-фитохелатин» в вакуоли происходит с помощью АВС-переносчика. Появляются данные, что в вакуоли могут аккумулироваться и другие тяжелые металлы, приносимые в нее с помощью других транспортных систем тонопласта. Было показано, что вакуоль является местом аккумуляции также Мо и Zn. Подобная компартментация Zn важна для выживания, например, растений ячменя. Более того, из растений арабидопсиса был изолирован ген (ТАТ), гомологичный гену переносчика цинка в клетках животных (ZnT). Перенос этого гена в растение и его активная экспрессия приводила к значительному увеличению устойчивости к цинку и аккумуляции этого металла в корнях при высокой концентрации его в среде. Следовательно, переносчик цинка мог участвовать в аккумуляции этого металла в вакуоли и тем самым в устойчивости растений (Кузнецов, 2005).
Таким образом, целый ряд механизмов повышения толерантности растений к тяжелым металлам направлен на удаление избыточного содержания металла из цитоплазмы и тем самым на предотвращение его возможных токсических эффектов. Для развития устойчивости к данному конкретному металлу используется не один, а несколько различных механизмов. Какого-либо единственного механизма, обеспечивающего толерантность растения к нескольким разным тяжелым металлам, не существует. Механизмы восприятия растением сигнала от действия тяжелого металла и трансдукции (передачи) этого сигнала к генам в настоящее время не известны.
Газоустойчивость растений
По характеру реакции у растений различают газочувствительность (т.е. скорость и степень проявления патологических процессов под влиянием газов) и газоустойчивость.
Газоустойчивость – это способность растений сохранять жизнедеятельность в присутствии в атмосфере вредных газов.
К ним относятся газообразные соединения: сернистый газ (SO2), оксиды азота (NO, NO2), угарный газ (СО), соединения фтора и др., углеводороды, пары кислот (серной, сернистой, азотной, соляной), фенола и др., твердые частицы сажи, золы, пыли, содержащие токсические оксиды свинца, селена, цинка и т.д.
Загрязняющие атмосферный воздух компоненты (эксгалаты) по величине частиц, скорости оседания под действием силы тяжести и электромагнитному спектру подразделяют на пыль, пары, туманы и дым.
Газы и пары, легко проникая в ткани растений через устьица, могут непосредственно влиять па обмен веществ клеток, вступая в химические взаимодействия уже на уровне клеточных стенок и мембран. Пыль, оседая на поверхности растения, закупоривает устьица, что ухудшает газообмен листьев, затрудняет поглощение спета, нарушает водный режим.
Наиболее сильно газы воздействуют на процессы в листьях. Косвенный эффект загрязнения атмосферы проявляется через почву, где газы влияют на микрофлору, почвенный поглощающий комплекс и корни растений. Кислые газы и кислые дожди нарушают водный режим тканей, приводят к постоянному закислению цитоплазмы клеток, изменению работы транспортных систем мембран (плазмалеммы, хлоропластов), накоплению Са, Zn, Pb, Сu. В этих условиях интенсивность фотосинтеза снижается из-за нарушения мембран хлоропластов. Кроме того, на свету быстро разрушаются хлорофилл а и каротин, меньше – хлорофилл b и ксантофиллы.
Адаптация растений к действию газов. У растений в процессе эволюции не могла сформироваться устойчивость к вредным газам, так как современная флора формировалась в течение тысячелетий в условиях, при которых их мало содержалось в атмосферном воздухе. Поэтому способность противостоять повреждающему действию газов основывается на механизмах устойчивости их к другим неблагоприятным факторам.
Для газоустойчивости существенна способность растений 1) регулировать поступление токсических газов, 2) поддерживать буферность цитоплазмы и её ионный баланс, 3) осуществлять детоксикацию образующихся ядов.
Несмотря на успехи в исследовании газоустойчивости растений, единой и общепризнанной теории устойчивости не существует. Значительный вклад в разработку различных вопросов теории газоустойчивости растений внесли В. Крокер, Н.П. Красинский, Е.И. Князева, М.Д. Томас, Ю.3. Кулагин, Г.М. Илькун, В.С. Николаевский и др.
В зависимости от механизмов, ее определяющих, газоустойчивость классифицируется как биологическая, анатомо-морфологическая и физиолого-биохимическая.
Под биологической газоустойчивостью следует понимать зависимость устойчивости растений от биологических особенностей (фазы роста и развития, скорости роста, наличия критических периодов), систематического положения и географического происхождения, преадаптации, экологической пластичности, светолюбия и др. Биологические механизмы устойчивости связаны с межродовым и межвидовым разнообразием. Например, крестоцветные более устойчивы, чем бобовые, из бобовых фасоль более устойчива, чем клевер, соя и т.д. Древесные растения (вяз, жимолость, клен) менее устойчивы по отношению к хлору, фтору, закиси азота, чем травянистые. У цветковых повреждаемость листьев зависит даже от их положения на побеге. Культурным растениям свойственна большая чувствительность к загрязнению атмосферы по сравнению с дикими видами.
Анатомо-морфологическая устойчивость связана с особенностями строения растений. Она проявляется в зависимости газоустойчивости растений от некоторых особенностей в анатомо-морфологическом строении листьев, ответственных за интенсивность газообмена и, следовательно, за скорость поглощения токсичных газов. Скорость поглощения газов зависит от числа устьиц, динамики их движения (степени открытия) в течение суток, толщины кутикулы, эпидермиса, толщины губчатой ткани, отношения высоты палисадной ткани к высоте губчатой и объема полостей в губчатой паренхиме. Для устойчивых видов древесных и цветочных растений в отличие от неустойчивых характерны большее число устьиц на 1 мм2 поверхности листа; меньшая длительность и степень открытия их в течение дня; большая толщина кутикулы и наличие различных дополнительных покровных образований; меньшая толщина и вентилируемость губчатой ткани; меньшая величина отношения высоты палисадной ткани к высоте губчатой. Все эти признаки в строении обусловливают снижение газообмена и, следовательно, поглощение вредных, газов. Для газоустойчивых видов характерны признаки ксероморфных черт в строении листьев, а для неустойчивых видов – мезоморфных.
Физиолого-биохимическая устойчивость определяется индивидуальными особенностями метаболизма растений, скоростью протекания биохимических реакций, способностью утилизировать ядовитые вещества, связывать их белками цитоплазмы и т.д.
К физиологическим механизмам устойчивости можно отнести состояние покоя у растений, которое выработалось в ходе эволюции как приспособление к перенесению неблагоприятного периода года, характеризующегося низкими температурами или продолжительными засухами. Резкое снижение интенсивности газообмена при одновременном усилении развития покровных тканей обеспечивает зимующим побегам деревьев и кустарников высокую газоустойчивость.
Возрастание уровня сахаров, аскорбиновой кислоты, азотосодержащих веществ в листьях также способствуют повышению газоустойчивости. Поддержание ионного баланса и буферных свойств цитоплазмы может быть связано с уровнем в клетках катионов (К+, Na+, Са2+), способных нейтрализовать ангидриды кислот. Обычно растения, устойчивые к засухе, засолению и некоторым другим подобным воздействиям, имеют более высокую газоустойчивость, возможно благодаря способности регулировать водный режим и ионный состав.
Группы устойчивости растений. Обычно по степени устойчивости выделяют три категории растений: устойчивые, среднеустойчивые и неустойчивые. Данная классификация была дана В.С. Николаевским. Критерием устойчивости служит размер площади некрозов в процентах от общей поверхности листа. Кроме того, могут использоваться такие показатели, как уменьшение всхожести семян, энергия роста и урожайность растения, метод биологических тестов и некоторые физиолого-биохимические, анатомо-морфологические показатели.
Повышение устойчивости растений к загрязняющим газам. Способы повышения устойчивости растений к вредным загрязнениям могут быть различными.
Селекционные методы. Используя высокоустойчивые формы, следует создавать новые формы и сорта еще более устойчивых растений. Успешные работы по отбору и размножению высокоустойчивых к сернистому газу и фтору особей хвойных растений ведутся в Германии. При нахождении таких особей их затем размножают вегетативным путем с целью сохранения признака высокой устойчивости у новых растений. В США созданы устойчивые к фотохимическому смогу сорта сои, шпината, картофеля и томатов. Исследования по выведению устойчивых к фитотоксикантам растений разворачиваются ныне в разных странах.
Агротехнические приемы. На плодородных почвах растения меньше страдают от загрязненности воздуха и оказываются более долговечными. При внесении удобрений в почву нейтрализуются накапливающиеся в ней вредные вещества, улучшаются условия существования микроорганизмов, способствующих детоксикации и нейтрализации вредных примесей.
Важное значение для нейтрализации кислых газов, поступающих с осадками в почву, имеет известкование.
Газоустойчивость повышается также в результате предпосевной обработки семян слабыми растворами микроэлементов (марганца, кобальта).
Для повышения устойчивости растений к токсическим веществам следует правильно снабжать растения водой. Обычно чем лучше водоснабжение растений, тем шире открыты устьица, интенсивнее транспирация и более активное поступление токсикантов. Вместе с тем осадки смывают с листьев вредные вещества, вымывают из их тканей хлор, сернистый газ и т.д. Удаление пыли способствует усилению процессов жизнедеятельности листьев.
Важное значение в повышении устойчивости растений к фитотоксикантам имеет характер размещения растений в посадках. Отдельно стоящие деревья и кустарники более подвергаются их действию по сравнению с теми, которые находятся внутри древостоя. По этой причине посадки деревьев в зоне атмосферных загрязнений должны располагаться достаточно плотно.
Под влиянием загрязнителей происходит замедление роста побегов, более быстрое их старение. В этих условиях путем обрезки растений можно стимулировать процессы омоложения вегетативных и генеративных органов. Положительное значение в защите неустойчивых к фитотоксикантам растений имеет посадка перед ними устойчивых деревьев и кустарников.