

Тимин_О_А_Лекции_по_общей_биохимии
.pdfСтроениеобменнукл отидов |
160 |
|
|
|
|
Н А РОУБШМЕЕННИАЕ П И Р ИОМСИНДОИВНАО В Ы Х Н И Й
|
|
|
|
β -АМ И Н О И З О Б У Т И Р А Т А Ц И Д У Р ИЯ |
|
|
|
|||
|
Таккакпродуктыкатаболизмапиримидиновхорасошо, воримы |
|
|
|
онихорошов |
ы- |
||||
водятсямочойи |
|
клиническиесимптомы |
патологий слабовыражены. |
|
|
|
||||
|
Примеромп |
одобного нарушенияявляется β-аминоизобутиратацидурия, |
прикотором |
|||||||
имеется |
дефектсоответствующейтранса |
миназы.Заболеванпротекаетбессиимптомно |
|
|
|
|||||
встречается,основном,уяпонцевикитайцевоколо( 25%). |
|
|
|
|
|
|
|
|||
|
|
|
|
|
ОР О Т А Т А Ц И Д У Р И Я |
|
|
|
|
|
|
Оротатацидурия (или |
оранжевкристаллоурия |
) можетбытьнаследственнойиприо |
|
|
б- |
||||
ретенной. |
|
|
|
|
|
|
|
|
|
|
Наследственнаяформа |
|
|
|
|
|
|
|
|||
|
Наследственнаяформа |
оротатацидурии весьмаредка |
связанаодновременнымдефе |
|
к- |
|||||
том оротатфосфорибозилтрансферазы и оротидилдекарбоксилазы,таккакэтиферменты |
|
|||||||||
являюдвумяактцентрамиивнымисяодногобелка.Заболеваниепроявляется |
|
|
|
навторомп |
о- |
|||||
лугодиижизн |
гипохромной мегалобластическойанемией, |
лейкопенией, |
отставанием ум- |
|||||||
ственногоифизического |
развития.Д иагнозставприобнаружениитсявохлаждмоч нной |
|
|
|
|
|||||
оранжевыхигольчакристаллоротовкислых.Боолезньвсложняетсяйтытем,что |
|
|
|
|
прин е- |
|||||
достатке УТФиЦТФактивируютсяпервыереакци |
|
исинтезаорот |
овойкислоты |
.Приотсу |
т- |
|||||
ствиилеченияг наступаетбельвпервыегодыжизни. |
|
|
|
|
|
|
|
|
||
|
Однпервопричиной,такак заболеванияявляется |
|
|
|
"пиримидиновыйголод |
|
",его |
|||
можнодостатуспешкорректироватьчнсп мощьюприемаурид, на |
|
|
|
|
этом серьезно |
|||||
неопасаясьпередозировки. |
|
|
|
|
|
|
|
|
||
Приобретеннаяформа |
|
|
|
|
|
|
|
|
||
|
Эта болеечастая |
форма оротатацидурииможетнаблюдаться: |
|
|
|
|
|
|||
o |
придефектекаких |
|
-либоферментовсинтезамоче,кромекарбамоилины |
|
|
|
фосфат- |
|||
|
синтетазы.Приэтомкарбамоилфосфатмитохондрийвнорме( |
|
|
|
используемыйдля |
обра- |
||||
|
зования мочевины)выходитизиспоизбыточнльзуетсясинтезаоротоговой |
|
|
|
|
|
||||
|
кислоты.Заб бычнолевансопргивождаетсяпераммониемией. |
|
|
|
|
|
|
|||
o |
прилеченииподагрыаллопуринолом,которыйпревоксипуринолмононуращается |
|
|
|
|
к- |
||||
|
леотид,являющийсяингибитор |
|
оморотат декарб,чтоксилазыпятьжеведетнакопл |
|
|
е- |
||||
|
ниюоротата. |
|
|
|
|
|
|
|
|
|
Н У КИЛНЕУОКЛЗЕИОД ТЫ |
|
|
ИКДАЫК |
|
|
Л Е К АПРРСЕТПВАЕРН Н Ы Е |
|
А Т Ы |
|
||
Азотистоеснование |
аллопуринол испоприлеченииьзуетсяподагры,прмочек |
|
|
а- |
|
меннойболезнисиндромеЛеша |
-Нихана. |
|
|
|
|
Лечебныйэффектряда |
антивирусныхпрепаратов |
оснихованстроении |
– ония в- |
||
ляютсяпроиазотистыхводоснованийымиуклеозидов.Механизмдейстразл чныхя |
|
|
|
|
|
нуклеозидовос ихфосфорилированиизараженныхвирусомклеткахпревращ |
|
|
|
е- |
|
ниемвнуклеотиды, |
в конкуренциисобы |
чныминуклеотидамизавстраивание |
|
ввирусные |
|
нуклеинкислотыве |
,чтоостанаразмножвирусалива. ениет |
|
|
|
|
МАБИОСИНТЕЗЫР Ч Е
Основнойфигуройматричныхбиосиявляютсянуклеитезовкислоты.Опрен в е |
|
|
д- |
||
ставляютсобойполимерныемоле,всоставкоторыхулывходятазотистыеоснованияпяти |
|
|
|
||
типов,пендвухтиповозыостаткифосфорнойкислоты. |
|
|
|
||
Азотистыеснованиянуклеиновыхкисл |
|
отахмогутбытьпуриновымиаденин( ,гу |
а- |
||
н)ипиримидиновымицитоз( ,урацил,тимин). |
|
|
|
||
Взависимостиотстроенияуглеводавыделяютрибонуклеиновыекислоты |
|
– содержат |
|||
рибозу( |
РНК),идезоксирибонуклеиновыекислоты |
– содержатзоксирибозу( |
ДНК). |
||
|
|
О С НПООВ Н О Й |
С Т У Л А Т |
М О ЛБЕИК ОЛУЯОРГНИО Й |
И |
Вподавболслучаевяющемьшинстве |
|
передача наследственнойинформацииотмат |
е- |
||
ринскдочернейклеткиосуществляетсяприпомощиДНК |
|
(репликация).Для использо- |
|||
вания генетическойинформациисамойклеткойнеобходимыРНК,образуем |
|
|
ыенаматрице |
||
ДНК (транскрипция).ДалРНКнепосредственноучаствуютнавсехэтапахсинтезабелк |
|
о- |
|||
выхмолекул |
(трансляция),обеспечивающихструктурудеятельностьклетки. |
|
|||
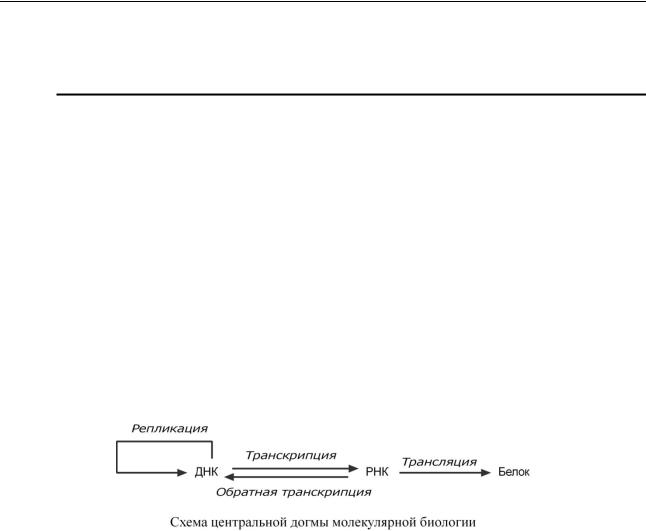
Навышесказоснованнома |
центральнаядогмамолекубио огииярной |
,согласно |
которойпереносгенетическойинформацииосуществляетсятолько |
|
от нуклеиновойкислоты |
(ДНКиРНК)Получателинформации. можбытьдругаяенуклеиноваякислотаДНК(или |
|
|
РНК)ибелок. |
|
|
|
|
|
С Т РНОУ КЕЛНЕИИЕН О В Ы Х |
К И С Л О Т |
|
|
СТ РДЕОЗОКСИРИБОНИ Е |
УКЛЕИНОВОЙКИСЛОТЫ |
|
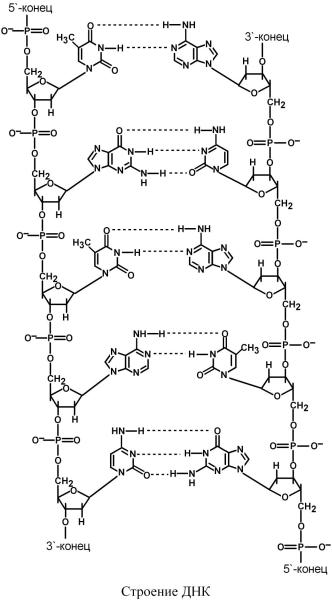
ДНКдезоксирибонуклеиновая( кислота)представляетсобойдвухцепочечнуюмолек |
|
||
лу,вкаждцепимономерамийвыступаютдезоксирибонуклеотиды |
|
|
|
dТМФ.Нуклеотисвязанымеждусобойчерезфосфаымосмеждутиные3'атомами5'ки |
|
|
|
углерососедна |
ихдезоксирибоз. |
СахарофосфатостовДНКзаряженый |
|
благодазарядуфосфатныхгрупп.Втояжевремяпуриновыепиримидиновыеоснования |
|
|
|
гидрофобны. |
|
|
|
ЦепиДНКоблнадаютправл,.е.именностьюют |
|
3'-конеци5 |
|
лельны,т.. |
напврасторонызныевлены |
|
.Имеетсякомплемеазотистыхосноварность |
ний:аденинкомплементарентимину( |
А=Т |
), гуанинкомплементаренцитозину( |
|
у-
– dАМФ,ГМФd,ЦМФd,
отрицательно
'-конец, цепиантипара л- а-
Г≡Ц) .
Матричныебиосинтезы |
162 |
|
|
|
|
СТ РХРОМЕ Н И ЕАТИНА |
|
|
|
|
ДНК – наиболееважнаячастьхромосом:дведвухцепмолекулыДНКбразуютчечные |
|
|
|
|
||
однухромосому.Наиболеехорошохромосомывидныпередмитвремязом |
|
|
|
него.Вп |
о- |
|
коящихсяклеткаххроматериалосомныйвыглядитнечеткорасповсемуряд.Вруелен |
|
|
|
|
|
|
такомсо |
стополучилянииназваниехроматин" "В.составехроматинавыделяют60%белка |
|
|
|
|
|
(гистоныкислыебелки), ДНК35%около5%РНК. |
|
|
|
|
|
|
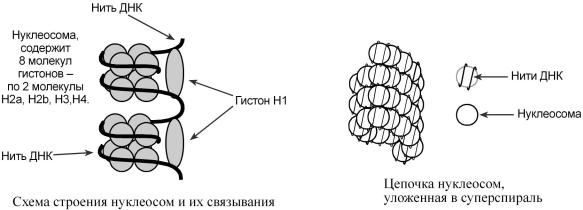
Хроматинуложеввидесф частицрических |
|
– нукл,соеосомдиненныхдругом |
|
|
|
|
нитьюД.НуклКпредставляетсобойомакомп |
|
лексучасткамолекулыДНКивосьми |
|
|
о- |
|
лекулгист.Вонуклставеновнахпоосм2дятсяомыгистоновлекулыН2 |
|
|
|
α,Н2 β,Н3,Н4. |
|
|
НитьДНК,последовательноконтакгисН2ируяонами |
|
α,Н2 β,Н4,Н3,Н3,Н4,Н2 |
|
β, Н2 |
α, |
|
наматываегистоновоеядро,комас"сяорое |
|
кирует" пар146основанийДНК.Гистон1 |
|
|
|
|
связывануклеоснаучасткетсявхивыходамойдаДНК,склеивая" "оборота2 маск" |
|
|
|
|
и- |
|
руя"еще20пароснований.Такимобразом,помощьюоднойнуклеосомызамаскировано |
|
|
|
|
||
166пароснований. |
|
|
|
|
|
|
Кроменуклеосом,вядпр |
исутствуютеще2 |
уровняукладкиДНК |
:фибриллыдиаме |
т- |
||
ромнм10,состоящцепочкииз нуклеосом,волок,диа30,обмаетразующиесяом |
|
|
|
|
||
призакручиваниифибрилл |
суперспираль.Навиток |
такой спириходитсяали6 |
-7нукл |
е- |
||
biokhimija.ru |
ТиминО.А. |
Лекции по общей биохимии (2018г) |
163 |
|
|
|
|
осом.УчастокДНКмеждунуклеосомаминазывспейсерным(етсянгл: |
|
|
space – простран- |
ство),егодлинаварьируетзависимостиотвидаорганизматипаклетчеловека.У она |
|
|
|
составоко50парнуклеотидовояет. |
|
|
|
Благодаряналичиюнуклеосомдостигаетсяуменьше |
|
ниеразмеровхромосомы |
враз7, |
далее происходитукладкав |
суперспираль фибриллы исуперсуперспираль„ " |
волокна. |
|
|
|
СТ РРИБОНУКЛЕИНОЕ Н И Е |
ВЫ Х КИСЛОТ |
|
|
|
РибонуклеиноваякислотаРНК() |
представляетсобойпоследовательностьрибонукле |
|
о- |
|||
зидмонофосфатов – АМФ,ГМФ, |
ЦМФ,УМФ |
,связанныхдругдругом5 |
'-3'-фосфо- |
|||
диэфирнымисвязями. |
Построению |
РНКотличаетсяДНКоднонитевойструктурой, |
|
значи- |
||
тельмемноьшейлекулярноймассой, |
|
наличиурацвмтиестомиларивместобозына |
|
|
||
дезоксирибозы. |
|
|
|
|
|
|
Вклеткеприсучетырествует |
|
типаРНК: |
|
|
|
|
МатричныеРНК |
(мРНК)представляютсобойлинейнуюпоследовательностьнукле |
|
о- |
|||
тидов.К5 |
'-концумолекулыприсоединенметилгу |
|
анозиндифосфат,на3 |
'-концеимеетсяп |
о- |
|
лиадениловаяпоследовательность |
|
(смниже) |
.Ихфункция |
– информаци,т.е.переноснная |
|
||
информацииструкбелДНКткуремеовихсинтезату. |
|
|
|
|
|
|
|
МалыеРНК |
исподсозреванияль мРНКуютсянекоторыхдругихклеточныхпр |
|
|
о- |
|||
цессов. |
|
|
|
|
|
|
|
РибосомальныеРНК |
(рРНК)прокариэук риотзличныличаютсявеличиной |
|
|
|
|||
седиментацск( оростседанмолекуиия |
|
|
|
лыприцентрифугировании). |
Ониучаствуютп |
о- |
|
строенииибосом. |
|
|
|
|
|
|
|
У прокариот есть тразновидностирРНК: 5 |
S, 16S и23 S. Малую(30S)субчастицу |
|
|||||
рибосом образуютбелк |
|
ии 16S-рРНК;большую(50S) |
субъединицу – белки, 23S-рРНКи |
||||
5S-рРНК. У эукариот – всоставер |
ибосом четыреразновидности |
рРНК: 5S, 5,8S, 18S и28 S. |
|||||
Малую(40S)субчастицуобразуютбелки18S |
|
|
|
-рРНК,большую(60S) |
– белки5S |
-, 5,8S-, |
|
28S-рРНК. |
|
|
|
|
|
|
|
Матричныебиосинтезы |
164 |
|
|
|
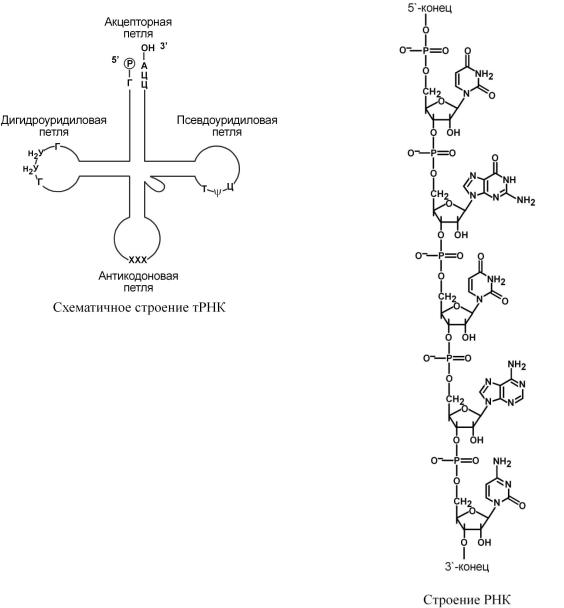
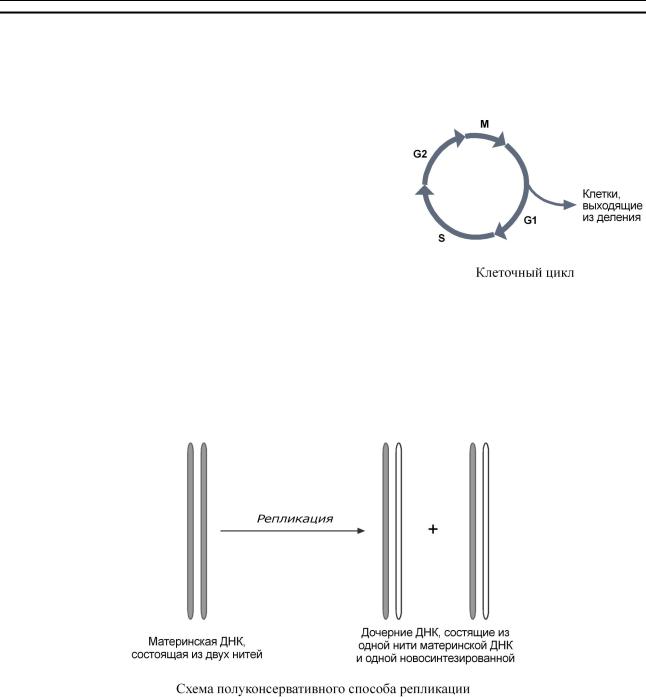
ТранспортныеРНК |
(тРНК)бактерийэукариотвключают73 |
-93нуклеотида.Онип |
е- |
||
реносятаминокислизцитозоляриб. сомамты |
|
На5 |
'-концетРНКнаходитсягуаниловый |
|
||
нуклеотид,на3 |
'-конце – триплетЦ |
-Ц-А. ВторичнаяструктуратРНКнапоминаетклеверный |
|
|||
лист,атретичная |
– латинскуюбукву |
L.В "клеверномлисте |
" выделяютчетыреучасткаили( |
|
||
ветви,петли),каждыйизкоторыхимеетсобственнуюфункци |
|
|
: |
|
||
o |
антикодоновый – соединяетсякодономматричнойРНКврибосоме, |
|
|
|||
o |
псевдоуридиловый – отвечаетзасвязываниерибосомой |
, |
|
|||
o |
дигидроуридиловый – отвечаетзасвязывание |
аминоацил-тРНК-синтазой, |
|
|||
o |
акцепторный – связываетпереносимуюаминокислоту |
. |
|
|||
biokhimija.ru ТиминО.А. Лекции по общей биохимии (2018г) 165
|
|
|
С И НТУЕКЗКЛИЕ И Н О В Ы Х |
С Л О Т |
|
|||
|
|
|
|
Р Е П Л И К А Ц И Я Д Н К |
|
|
||
|
СинтезДНКпроисходитнебеспорядочно,ав |
|
|
|
|
|||
строгоопределепержклеткизод.Всегонныйи |
|
|
|
|
|
|||
выделяютфазы4 |
|
клеточногоцикла |
:митозМ(), |
син- |
|
|
||
тетическую( |
S),пресинтетическую( |
G1, отангл. |
gap – |
|
|
|||
интервал),постсинтетическую( |
G2). |
|
|
|
|
|||
|
Важноеучастиеврегуляциисменыфазклето |
циклины |
|
ч- |
|
|
||
ногоциклазанимают |
|
– белкимассой |
|
|
|
|||
35-90 кДа,уровенькоторых |
меняетсявходекл то |
ч- |
|
|
||||
ногоцикла |
.Пофункциициклин |
ы – этоактиваторные |
|
|
||||
субъферментовдиницы |
циклин-зависимыхкиназ |
|
|
|
||||
(ЦЗК) |
.Активныекомплексыциклин |
|
-ЦЗКфосфор |
и- |
|
|
||
лируютвнутрбелки, клеточныезменяяихактивность.Этим |
|
|
|
обеспечиваетсяпродвижение |
|
|||
поклеточномуциклу. |
|
|
|
|
|
|
||
|
Синтезрепликация( ,удвоение)ДНКпроисх |
|
|
одитв |
S-фазуклеточногоцикла |
,когда |
||
клетготовитсякделениюа. |
Механизмреплик,какустановМэтьюцииМезельсонили |
|
|
|||||
ФранкСтав1957льин |
|
|
г, полуконсервативный, т.е.накаждойнитиматерДНКсинской |
н- |
||||
тезирудочернкопи. тсяя |
|
|
|
|
|
|
||
тов: |
Как любой матричныйбиосинт,репликациятребуналичиязсколькихт |
компонен- |
||||
матрица – веероли выступаетматеринская |
нить ДНК, |
|||||
o |
||||||
o |
растущаяцепь |
– дочерняя нить ДНК, |
|
|||
o |
субстрадлясинтыеза |
– dАТФ, |
dГТФ,Цd, ТФ |
, |
||
o |
источникэнергии |
– dАТФ, |
dГТФ,Цd, ТФ |
, |
||
o |
ферменты. |
|
|
|
|
|
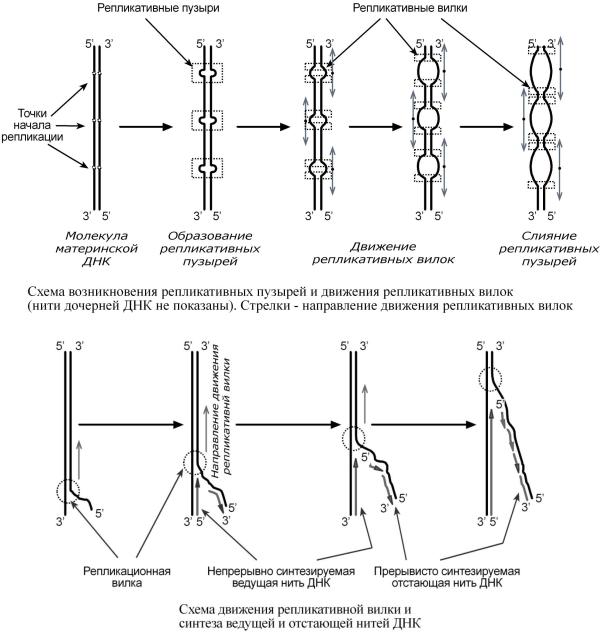
СиДНКначинатезвопручасткеделется,получившнаныхточказваниех |
ori |
||
(ан гл. |
origin – начало) |
. Нак ДНКждоймлекточекпитающих |
ori насчитываетсяоколо100. |
Репликация распространяетсяотэтихучавобековороныпонитДНКм |
собразова нием |
||
репликативных "пузырей". Вкаждтакпузыре"ом"имеютсядве |
репликативные "вил- |
||
ки",вкоторыхпроирасплетаниеходит,раскручиванепо редственныйДНК.тезие |
|
||
Приэтом |
епликатвилкудаляютсядругиотвныедруга.Вц всяломрепликацияДНКуэ |
у- |
|
кариотзаканчиваетсячасов9.
Матричныебиосинтезы |
|
|
|
|
|
|
|
166 |
|
|
|
|
|
|
|
||
СинтезновойцепиДНКидетвнаправленииот5' |
|
|
-конца |
3'-концу,т.е. 5' |
-конецновой |
|
||
ДНКостаетсясвободным,следнуклеотидыющиеприсоединяются3' |
|
|
|
-гидроксильной |
||||
группепредыдущегонуклеотида. |
|
|
|
→ 3' непрерывно (т.е.обычнымприсоедин |
|
|
||
Вреплвивкативнойлке |
направлении5' |
|
е- |
|||||
ниемпоследующнуклеотпредыдущимдовх |
|
черезС |
3 иС |
5) синтезируетсяолько |
|
одна |
||
нить,аименнота,длякоторойнаправлнаправлениемсинтезасовп даетдвижения |
|
|
|
|
|
е- |
||
пликативнойвилкисоответствуе |
|
тнапрмавлентеринскойнит ю |
|
3' → 5'Помере. распл |
е- |
|||
танияДНКдвиженреплинакативнойэтойлкияматеринскойни крываютсяучас |
|
|
|
|
т- |
|||
ки,гдевозможно |
безостановочное удлинение ведущей дочерней нити. |
|
|
|||||
Направление5' |
→ 3'для |
другой дочернейнитиДНКпроти |
|
вополождвижениюрепло |
|
и- |
||
катви.Поэтвнойлкис этойнтезмутстающейнитивнаправлении( 5' |
|
|
|
→ 3')возможен |
||||
толькопослерасплетаниячастиДНКосвобожденияучасткадлясинтеза. |
|
|
|
|
|
|
||
Такимобраз,синтездочернейДНКмидетфрагментарно.Поименияпонисскл го |
|
|
|
|
е- |
|||
дователясинтезируемыена |
|
отстающей цепиотрезкиДНКназвали |
|
фрагментыОказаки. |
|
|
||
biokhimija.ru |
|
ТиминО.А. |
|
Лекции по общей биохимии (2018г) |
|
167 |
||||||
|
|
|
|
|
|
|
|
|
||||
|
В хронолпорядкесобытиягичрепликацразвескомритываютсяследуимерно |
|
|
|
|
|
|
ю- |
||||
щимобразом: |
|
|
|
|
|
|
|
|
|
|
|
|
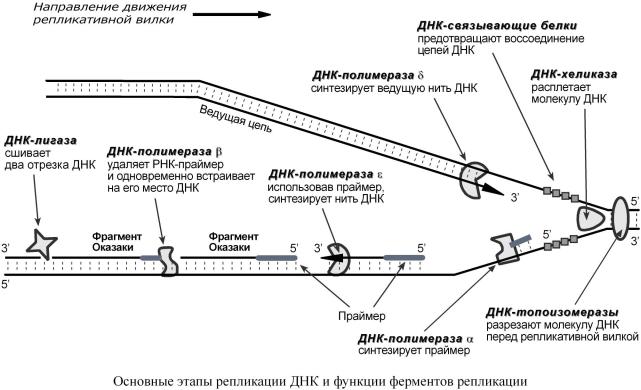
1. |
ДНК-топоизомеразы,находясьпередреплви,ркативнойлкой |
|
азрезают молекулу ДНК |
|||||||||
|
дляоблегченияеерасплетанияраскручива |
|
|
|
|
|
ния. |
|
|
|
|
|
2. |
ДНК-хеликазы,следуятопоизомеразами |
|
|
, раскручивают ирасплетают молекулу ДНК. |
||||||||
3. |
ДНК-связывающиебелки |
|
(ДСБ) |
связывают расплетенные нити ДНК истабилизируют |
||||||||
|
их,недопуская |
обратного "слипания" другсдругом. |
|
|
|
|
||||||
4. |
ДНК-полимераза δ (греч.: |
δ – дельта),с |
|
оглассоскодвиженваноростьюрепликатия |
|
|
в- |
|||||
|
нойвилки , осуществляетэлонгацию |
|
ведущей цепидочернейДНК |
|
внаправлении 5'→ 3' |
|||||||
|
наматрице одизнитейой |
|
материнскойДНК |
(скорость100парнуклеотидов |
|
секунду) |
. |
|||||
5. |
ДНК-полимераза α присоединяетсяк |
другой нитиДНКсразупослерасплетания |
|
|
в |
|||||||
|
направлении 5'→ 3' синтезирует праймер (РНК-затравку) |
– последовательностьРНК |
на |
|||||||||
|
матрице ДНК длинот10до200нуклеотидовй |
|
|
. Послеэтогоферментудаляется |
|
нити |
||||||
|
ДНК. |
|
|
|
|
|
|
|
|
|
|
|
6. |
ДНК-полимераза ε проводитс |
интез фрагмента (длина150 |
-200нуклеотидов) |
отстающей |
||||||||
|
цепидочернейДНК |
|
впродолжениепраймера |
|
|
. Онарабдтехопортает,поканевстретит |
|
|
|
|||
|
праймер предыдущегофрагмеОказакиси( нтезированного |
|
|
ранее). Послеэтогод |
анный |
|||||||
|
ферментудаляетсяцепи. |
|
|
|
|
|
|
|
|
|
|
|
7. |
ДНК-полимераза β встает вместо ДНК-полимеразы ε,движется |
томже |
направлении |
|||||||||
|
(5'→ 3') иудаляет рибонуклеотидыпраймера |
|
,одновременновстраивая |
дезоксирибонук- |
||||||||
|
леотиды наихместо.Ферментрабдполноготаетудаленияпраймера,послечего |
|
|
|
|
|
схо- |
|||||
|
дит сцепи. |
|
|
|
|
|
|
|
|
|
|
|
8. |
ДНК-лигаза производитс |
шивкуотрезк |
а, |
синтезированного ДНК-полимеразой ε, иучас |
т- |
|||||||
|
ка отстающейцепидочернейДНК |
|
|
|
, встроенного ДНК-полимеразой β, (фрагментовОк |
а- |
||||||
|
заки). |
|
|
|
|
|
|
|
|
|
|
|
Матричныебиосинтезы |
|
|
|
|
|
|
|
|
168 |
|||
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
РЕ Г УРЕПЛЯ Ц ИКАЦИИЯ |
|
|
|
|
|
||
|
|
|
|
|
Лекарственнаярегуляция |
|
|
|
|
|
||
|
1. Дауномицин и актиномицин D интеркалируют,..встраиваютсямеждунитей |
|
|
|
||||||||
ДНКипрепятствуютродвижфе меплен. итовкациию |
|
|
|
|
|
|
|
|||||
|
2. Новобиоцин снижаетактивностьДНК |
|
-гируба,зыктеподавляяихрийазмножение |
|
|
. |
||||||
|
3. Мелфалан и циклофосфамид преворганизмеращаютсявсоединения,которыев |
|
|
|
ы- |
|||||||
зываютприсоединениегруппы |
CH3 или C2H5 катому |
N7 |
гуанозина обеихцепмочлекулы |
|
|
|||||||
ДНК,чтоприводиткперекрестнойсшивкедвухспиралейДНКостановкерепликации. |
|
|
|
|
|
|
|
|||||
|
|
|
|
|
П О В РИРЕ ЕЖПДАЕРНАИЦЯ |
|
И Я Д Н К |
|
|
|||
|
Таккакнагеномлюбойнедеклпостоянноеткиящейсяоказываетвлияниеокружа |
|
|
|
|
|
|
ю- |
||||
щаясреда,товполневерповрежденияятнысоставенукл,такжезможнотидавстра |
|
|
|
|
|
|
и- |
|||||
ваниенеправильногонуклеплиотида.Такиенарушенияациибыс |
|
|
|
|
троопределяются |
|||||||
специальнымифермент,пораженныйучудаляетсямисток |
|
|
|
экзонуклеазами,заполняется |
||||||||
ДНК-полимеразой β исшивается |
ДНК-лигазой. |
|
|
|
|
|
||||||
|
Вслучаеизмструктурыненияоснованиянапример( ,егодезаминирование)этоосн |
|
|
|
|
|
о- |
|||||
ваниеудаляется |
|
|
ДНК-N-гликозидазой,затемдругиферудаляетментамидезоксяирибоза |
|
|
|
||||||
инаееместо |
|
ДНК-полимеразой β и ДНК-лигазой встраиваетсянужныйнуклеотид. |
|
|
|
|||||||
|
|
|
|
Г И Б Р И Д И З А Ц И Я Д Н К - Д Н К И Д Н К - Р Н К |
|
|
||||||
|
ЕслинагрерастворДНКвышеьтемпературы90 |
|
|
°СилисдвинутьрНрезкощелочную |
|
|
||||||
илирезкокислую |
|
|
сторводородные, связимеждуДНКтямиразрушаются |
|
|
|
и двойная |
|||||
спиральрасплетается.Происходит |
|
денатурация ДНКили,по |
|
-другому, |
плавление.Еслиуд |
а- |
||||||
литьагрессивныйфакт,топроисходит |
|
|
ренатурация или отжиг.ПриотжигенитиДНК |
|
|
|||||||
"отыскивают" |
комплементарныеучасткидруг |
|
другаи,вконцеконцов,вновьсворачиваю |
|
|
т- |
||||||
сявдвойнуюспираль. |
|
|
|
|
|
|
|
|
|
|
|
|
|
Есливоднойпробирке" "провесплавлениетсмесижигДНКчеловекаимыши,то |
|
|
|
|
|
|
|
||||
некоторыеучасткицепейДНКмышибуд |
|
|
|
|
утвоссоединятьсякомплементарнымиучастк |
|
|
ами |
||||
цепейДНКчеловекасобразованием |
|
гибридов.Числотакихучастковзависитотстепени |
|
|
|
|||||||
родствавидов.Чемблвидымеждужесобой,тембольшеучастковкомплементарности |
|
|
|
|
|
и- |
||||||
тейДНК.Это |
|
явление называется гибридизацияДНК |
-ДНК. |
|
|
|
||||||
|
ЕсливраствореприсутствуетРН |
К,томожносуществить |
гибридизациюДНК |
-РНК. |
||||||||
Такаягибридизация |
|
помогаетустановитьблизостьопредепослеДНКнныхдовательностей |
|
|
|
|
|
|||||
скакой -либоРНК. |
|
|
|
|
|
|
|
|
|
|||
|
ГибридизацияДНК |
-ДНКиДНК |
-РНКиспользуетсякакэффективнсредствмол ое |
|
|
|
е- |
|||||
кулярнойгенетике. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
С И Н Т Е З Р Н К – Т Р А Н С К Р И П Ц И Я |
|
|
|
||||
|
Прчемначнутждесинтезирбелки,информациюихватьсябстроениинеобходимо |
|
|
|
информацион- |
|||||||
"достать"изДНКидоставитьеекместусинтезабелков.Этимзанимаются |
|
|
|
|
||||||||
ные или матричные РНК.Одновременнокленужныткеранспортерыаминокислот |
|
|
|
|
– |
|||||||
транспортные РНКиструктукомпонентыр,гансинтезирующихныебелоклл, |
|
|
|
– рибосо- |
||||||||
мальные РНК. |
ВсяинформстроенанцирибосомальныхспоряиРНКтныхакже |
|
|
|
|
|
||||||
находитсявДНК. |
|
|
|
|
|
|
|
транскрипции дансДНКныха |
|
|
||
|
Поэтомусуществуетпроцесспереписыванияили |
|
|
|
|
|
||||||
РНК (англ. |
transcription – переписывание) |
– биосинРНКнамаДНКт.рицеез |
|
|
|
|
||||||
тов: |
Каквлюбомматричномбиосинтезевтранскрипциивыднеобходимых5 ляютэлеме |
|
|
|
|
|
|
н- |
||||
матрица – однаизцепейДНК |
, |
|
|
|
|
|
|
|||||
o |
|
|
|
|
|
|
||||||
o |
растущаяцепь |
|
– РНК, |
|
|
|
|
|
|
|
||
biokhimija.ru |
ТиминО.А. |
Лекции по общей биохимии (2018г) |
169 |
|
|
|
|
|
|
o |
субдлясинтезатрат |
– рибонуклеотидыУТФ(,ГТФ,ЦТФ,АТФ) |
, |
|
o |
источникэнергии |
– УТФ,ГТФ,ЦТФ,АТФ |
. |
|
o |
ферментыРНК |
-полимеразы ибелковыефакторытранскрипции |
. |
|
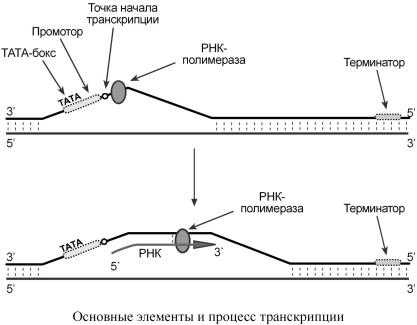
БиосинтезРНКпроисходитвучасткеДНК,которназываетсяй |
|
|
транскриптон,со |
д- |
||||||
ногокраяонграничен |
|
промотором (начало),сдругого |
– терминатором (конец). |
|
|
|||||
РНК-полимеразы эукариимеюподвеботсубъединицыльшихнесколькомалых |
|
|
|
|
|
|||||
субъединиц. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
СТ АТРАНД И И |
СКРИПЦИИ |
|
|
|
|
|
Выделяюттристадиитранскрипции: нициация,элонгацтерм. инация |
|
|
|
|
|
|
||||
|
|
|
|
Инициация |
|
|
|
|
||
Промоторсодержитстартовыйсигнал |
|
|
транскрипции – ТАТА-бокс – определенную по- |
|||||||
следоватнуклеотидовДНК, льность |
|
|
|
связывающую |
первыйфакторинициации |
|
|
|||
ТАТА-фактор.ЭтотТАТА |
-факторобеспечиваетприсоединение |
РНК-полимеразы ктой |
||||||||
нитиДНК,котораябудетиспользоватьсякачесшаблодляравенскрипции |
|
|
|
(матричная |
||||||
нитьДНК) |
.Таккак |
|
промоторассиметричен |
("ТАТА"),тоонсвязываетРНК |
-полимеразу |
|||||
тольководнориентации,чтойопределяетнаправлениетранскрипцииот5 |
|
|
|
'-конца3 |
'-концу |
|||||
(5'→ 3'). ДлясвязыванияРНК |
|
-полимеразыспромнеобходимтором |
|
ещеодинфактор |
|
и- |
||||
циации – σ-фактор (греч. |
σ – "сигма"),носразуп |
ослеинтезазатравочногофрагментаРНК |
|
|
|
|||||
(длиной8 |
-10рибонуклеотидов ) σ-фактотрываетсяфермента. |
|
|
|
|
|
||||
ДругиефакторыинициацраскручиваютспДНКпередральРНК |
|
|
|
|
-полимеразой. |
|
||||
Белковыефакторы |
|
|
Элонгация |
|
|
|
|
|||
|
|
элонгацииобеспечиваю |
|
тпродвижениеРНК |
-полимеразывдоль |
|
||||
ДНКирасплетают |
|
молекулу ДНКнапротяжениипримернонуклеотидных17 пар. |
|
|
|
|
||||
РНК-полимеразапродвигаетсясоскоростью40 |
|
|
-50нуклвсевкунотинапрадовулении |
|
|
|
||||
5'→ 3'. Ферменти |
спользует АТФ,ГТФ,ЦТФ,УТФодновременно |
|
вкач |
ествесубстратаи |
|
в |
||||
качестве источникаэнергии. |
|
|
|
|
|
|
|
|
||