

Тимин_О_А_Лекции_по_общей_биохимии
.pdfМатричныебиосинтезы |
|
|
|
|
|
|
170 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Терминация |
|
|
|
|
РНК-полимеразаостан,когддостигнетвитсятерминируюкодонов.Спомощьюих |
ρ-фактора (греч. |
|
|
|||||
белковогофактора |
|
терминации,такназываемого |
ρ – "ро"),матрицы |
|
||||
ДНКотделяютсяферментисинтезирмолекулаРНК, являетсяваннаяторая |
|
|
|
первичным |
||||
транскриптом,предшественникоммРНКилитРНКрРНКли. |
|
|
|
|
||||
|
|
|
|
П Р О Ц Е С С И Н Г Р Н К |
|
|
|
|
СразупослесинтезарвичныетранскриптыРНКпоразнымпричинамещенеимеют |
|
|
|
|
||||
активности, |
являются "незрелыми " ивдальнпретерпрядйшемизм,которыеваютнений |
|
|
|
||||
называются процессинг.У |
эукарипроцессингудтввидысеергаютсяпре |
|
|
-РНК,упрок |
а- |
|||
риот – толькопредшественникирРНКтРНК. |
|
|
|
|
|
|||
|
|
|
ПР О ЦПРЕДШС С И НЕСТВЕГ |
ННИКАМ |
РНК |
|
|
|
Притранскрипции |
участковДНК,несущихинформациюбелках,образуются |
|
|
|
гетеро- |
|||
генные ядерныеРНК,поразменамногопревосходящиеумРНК.Делотом,чтоиз |
|
|
|
-зам о- |
||||
заичнойструктурыгеновэтигетерогенинформативныеРНКвключаютсебя ( |
|
|
экзоны) |
|||||
инеинформативные( |
|
интроны)у частки. |
|
|
|
|
||
1. Сплайсинг (англ. |
splice - склеиватьвстык) |
– особыйпроцесс, |
|
в котором приучастии |
||||
малыхядерныхРНК |
|
|
происходинтроновудалениесохранениеэкзонов. |
|
|
|
|
|
2. Кэпирование (англ. |
cap – шапка) |
– происходитещевовремятранскрипции |
|
.Процесс |
|
состоитв |
присоединении |
к 5 '-трифосфатуконцевогоуклеотидапре |
-мРНК5 |
'-углерода |
|
biokhimija.ru |
|
ТиминО.А. |
Лекции по общей биохимии (2018г) |
|
171 |
|||||
|
|
|
|
|
|
|||||
N7-метил-гуанозина.Кэп"необходимдлящитымолекулыРНКотэкзонуклеаз |
|
|
|
|
,работа ю- |
|||||
щихс5 |
'-конца,атакжедлясвязываниямРНКрибосомой |
|
|
для началатрансляции. |
|
|
||||
3. Полиаденилирование – припомощиполиаденилат |
|
-полсиспользованиеммеразы |
|
|||||||
молекулАТФпроисходитприсоединениек3 |
|
|
'-концу РНКот |
100 до 200аденуклеиловых |
о- |
|||||
тид,формирующихвполи |
(А) -хвост. |
Поли (А) -хвостнеобходимдлязащитымолекулыРНК |
|
|
||||||
отэкзонуклеаз, |
работающихс3 |
'-конца. |
|
|
|
|
|
|||
|
|
|
ПР О ЦПРЕДШС С И НЕСТВЕГ |
ННИКАР |
РНК |
|
|
|||
ПредшественникирРНКявляютсяболеекрупнымимолекуламипосравнениюсозр |
|
|
|
|
|
е- |
||||
лымирРНК |
.Ихсозревсводитсяк аниезрпреризаниюРНКнабомелкиелеесомной |
|
|
|
|
|
|
|||
формы,которыеуженепосредственноучаствуют |
|
|
|
вформированиирибосомы. |
Уэукариот |
|||||
существуют 5S-, 5,8S-, |
18S-,и28 S-рРНК.Приэтом5 |
S-рРНКсинтезируетсяотдельно,а |
|
|
||||||
большаяпрерибосомная45 |
S-РНКрасщепляетсяспецифичнынуклеазаобразованиемми |
|
|
|
|
|||||
5,8S-рРНК, 18 |
S-рРНК,и28 |
S-рРНК. |
|
|
|
|
|
|
||
У прокариотмолекулы |
рибоРНКсоиныемальнвсемпосвоим(5Sйствам |
|
|
|
-, 16S- |
|||||
, 23S-рРНК),чтоявляетсяосновойизобретенияиспользованияряда |
|
|
|
|
антибиотиковмед |
и- |
||||
цине. |
|
|
|
|
|
|
|
|
|
|
|
|
|
ПРОЦЕССИНГПРЕДШ ЕСТВЕ |
ННИКАТ |
РНК |
|
|
|||
1. Формирование на3 '-конце последовательности Ц-Ц-А.Дляэтуоднихго |
|
|
||||||||
пре-тРНКс3 |
'-концаудаляютсялишниенуклдо отиды |
|
"обнажения" триплетаЦ |
-Ц-А,удр |
у- |
|||||
гиприсоединениедетхэтойпоследовательности. |
|
|
|
|
|
|
|
|||
2. Формированиеантикодоновойпетли |
|
проипутемсплайсингаходитудаления |
|
н- |
||||||
тронавсреднчастипрей |
|
-тРНК. |
|
|
|
|
|
|
||
3. Модификация нуклеотидов вмолекуле |
путемдезам, инети, лирования |
|
|
|||||||
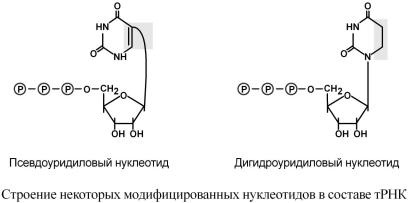
восстановлен.Наприм,образопсевриданиеяигидроуридина. |
|
|
|
|
|
|
|
|||
|
РЕГУЛЯЦИЯТРАНСКРИПЦИ |
И |
|
Таккактранскрипциясвязываетядро |
– "мозг"клетки,ееба"зн"анийбелки,раб" |
о- |
|
чихлошадок" |
клетки,тооткачесактивностиварнскрзависитобъемтехпциинтеза |
|
|
илииныхбелков,жизнедкл,ееспособностьткиятельнадаптик кружающейовться |
|
|
|
обстановке. |
|
|
|
Упрокариотэукариотрегуляциятранскрипциипроисходестественно, ,по |
|
-разному, |
|
хотянекоторыемоментыпохожи |
|
. |
|
Прокариоты
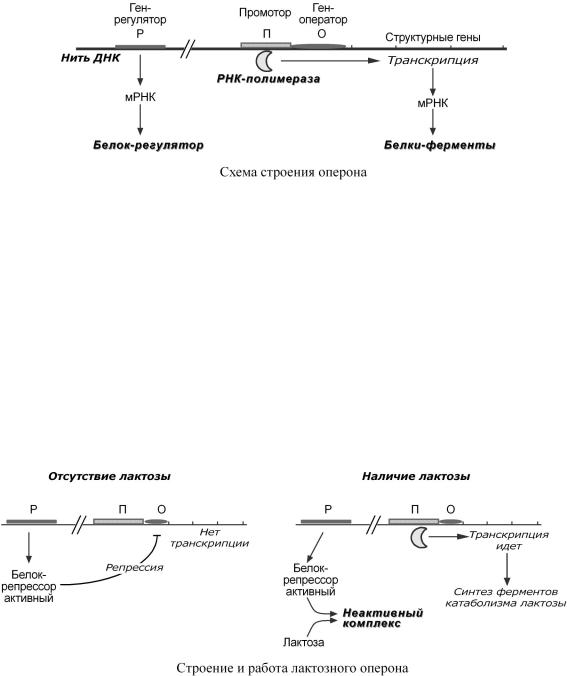
Регуляциябиосинтезабелкапр кариотсущестнауротранскрипциивляется мРНК.Внастоящеевремяпринятатеоперрия,сфонармулирФрансуаЖакованнаябом Жаком М.Воноснтеориилежатвеследующиепонятия:
Матричныебиосинтезы |
|
|
|
172 |
|
|
|
|
|
||
o |
оперон – группатесносвязанныхмеждусобойгенов,которыепрограммируютобр |
|
а- |
||
|
зованиеструктубелковиферклныхм,ентовтке |
|
|
|
|
o |
конституитивные гены – те, которые активны вклеткахвсегда,независимоотее |
к- |
|||
|
тивности, |
|
|
|
|
o |
индуцибельные гены – те,которые |
считываются при необходимости, |
|
||
o |
ген-регулятор – ген,регулирующийработу |
оперона, |
но невходящийегос .став |
Он |
|
|
синтезирует белок-регуляторчаще( называемый |
белок-репрессор),котможетбытьрый |
|
||
|
в активной или неактивной форме, |
|
|
|
|
o |
ген-оператор – учаДНК,стпосвязыватьсяксобныйбелком |
|
-регулятором,и" |
е- |
|
|
шающий"нужнорабРНКтать |
-полимеразеилинет. |
|
|
|
Лактозныйоперон |
|
|
|
|
|
|
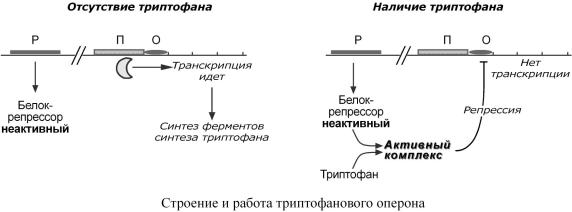
Лактозныйоперонвцеломотвечаетзакатаболизмлактозы. |
|
|
|
|
|
Приизучении |
E.coli былозамечено,что |
вклеткеможетбытьдве |
взаимоисключающие |
|
ситуации: |
|
низка,есливсреде |
имеется |
||
o |
активодногоизферментостькатаболизмалактозыв |
||||
|
глюкоза. |
|
повышается вобраси,т.е.нойуациипри |
|
от- |
o |
но активность этого ферезкомента |
|
|||
|
сутствииглюкозы |
ипри налактозыичии |
. |
|
меха- |
|
Наоснованииэтихнабл |
юденийбылапредложенасхемарегуляцииоперонапо |
|
||
ниндукциизму |
: |
|
|
|
|
1В. |
отсутствиелактозы |
активный белок-репрессор связываеопераиблокируетсяором |
|
|
|
синтезмРНК,кодирующейферментыкатаболизмалактозы.Врезультатеэтиферменты
необразуются .
2. Есглюкозынети,а |
лакестьоза |
,топоследняявязываетсябелком |
-репрессоромии |
н- |
гибируетего,недасваяязатьсягеном |
|
-оператором.ЭтозволяетРНК |
-полимеразе |
|
biokhimija.ru |
ТиминО.А. |
Лекции по общей биохимии (2018г) |
|
173 |
|||
|
|
|
|
|
|||
|
счинформтывать,отвечающуюз цию |
синтезферментов |
катаболизмалактозы,си |
н- |
|||
|
тезировать мРНК. |
|
|
|
|
|
|
|
Такимобразом, |
лактозаявляется |
индуктором транскрипции. |
|
|
||
Триптофаопероновый |
|
|
|
|
|
|
|
|
Триптофаоперовцеломотвечаетнзасинтезовыйтриптофана. |
|
|
|
|
|
|
|
Функцтриптофаонированиеоперонанекоторомсмыслеовогопротивоположнола |
|
|
|
к- |
||
тозному. Регуляцияосуществляется |
по механизмурепрессии |
|
. |
неактивном состоя- |
|||
1В. |
отличиелактозноперона,белокго |
-репрессорсинтез |
ируетсяв |
||||
|
нинеможетизаблокироватьрансгенов, одирующихрипциюферментысинтеза |
|
|
|
|
|
|
|
триптофана.Синтез |
этой аминокислотыбудетвклеткепродолжаться |
|
|
|
ехпор,покав |
|
|
питательной среденепоявтри. птофантся |
|
активирует его.Далеетакойактивный |
||||
2. |
Триптофансоединяетсябелком |
|
-репрессороми |
||||
|
комплексприсоединяетсягену |
-операиблокируетранскрипциюору.Такимобразом, |
|
|
|
||
|
приналичии |
триптофанавсреде |
прекращается еговнутрси,энтезклономяточный |
|
т- |
||
|
сяресурсыиэнергиябактериальнойклетки. |
|
|
|
|
|
|
|
Вэтомслучае |
триптофанявляется |
репрессором транскрипции. |
|
|
||
|
|
|
Эукариоты |
|
|
||
Внутрегуляцияиклеточная |
|
|
|
|
|
|
|
Существеусложэукариотическихнорганизмповлеклоноеениезасобпоявленией |
|
|
|
|
|
||
новыхспособоврегуляцииактивности |
|
|
транскрипции. |
|
|
||
Амплификация – этоувеличколичестваген,точовмиеногееопированиекратное |
|
|
|||||
одногогена.Есте,вполученныественнокопии |
|
|
|
|
равнозначныиодинаковоактивнобесп |
|
е- |
чиваюттранскрипцию. |
|
|
|
|
|
|
|
Энхансеры (англ. |
to enhance - усиливать) |
– это участкиДНКв10 |
-20пароснований, |
|
|||
способныезначительно |
|
усиливать экспрессиюгеновтойжеДНК.Вотличиепромоторов |
|
|
|||
онизначиудаленыоттрельно |
|
анскрипционногоучасткамогутрасполагатьсяотнегов |
|
|
|||
любомнаправлениик(5 |
|
'-концуили3 |
'-концу)Сами. энхансенекодируюткакиеы |
|
-либо |
||
белки,носпособнысвяз |
|
ыватьсярегулятбелкамип( одавляющрнымитранскр). имипцию |
|
|
|||
Сайленсеры (англ. |
silence – молчание) |
– участкиДНК,впрсхожинципеэнханс |
|
е- |
|||
рами,ноониспособны |
|
замедлять транскрипциюгенов,связываясьрегуляторнымибелк |
|
а- |
|||
ми (которыеееактивируют) |
|
. |
|
|
|
|
|
Пергеновстройка |
|
. Кподобнымпр цестноситсяам |
|
кроссинговер – обменучас |
т- |
||
камигомологичных |
хромосом, ибослееожныйпроцесс |
|
– сайт-специфичнаярекомбин |
а- |
|||
ция,котораяизменяетположениепорядокнуклпослеотивдгеноменыховат. льностей |
|
|
|
|
|
||
ПроцессингмРНК |
|
– некоторыепре |
-мРНКподвергаютсяразнымвариантамсплайси |
|
н- |
||
га( альтернсплайсингтивый |
)вре |
зультатечегообразуютсяразныемРНК,исоотве |
|
т- |
|||
ств,бесразнойннолкифункцией. |
|
|
|
|
|
|
|
Матричныебиосинтезы |
|
|
|
|
|
174 |
|
|
|
|
|
|
|||
ИзменениестабильностимРНК |
|
– чемвышепродолжительностьжизнимРНКвц |
|
о- |
|||
зокле,тембольшетки |
|
синтезируется соотвбелка.тствующего |
|
|
|||
Лекарственнаярегуляция |
|
|
Ингибирование |
|
|
||
|
|
|
|
|
|
||
1Гетер. социклическиеединения |
|
доксорубицин,дауномицин |
и актиномицин D об- |
||||
ладаютспособностью |
интеркалировать (встраиваться междунитей |
молекулы ДНК)между |
|
||||
двумясоседнимипарамиоснованийГ |
|
-Ц.Врезультатевозникаетпрепятствиедлядвижения |
|
|
|||
РНК-полимеразы( "заеданиемолнии |
")иострансанов. карипции |
|
|
|
|||
2. Рифампицин связывается |
β-субъединицейРНК |
-полимеразы прокариот иингиб |
и- |
||||
руе.Благодаряеттакойизбирательностидействиярифампициндействуеттольнабако |
|
|
|
|
е- |
||
риявляипредлтсяпаратомечениятуберку |
|
|
леза. |
|
|
|
|
3. α-Аманитин,октапептид |
бледнойпоганки( |
Amanita |
phalloides)блокирует |
||||
РНК-полимеразу II эукариот ипредотвращаетпродукциюмРНК. |
|
|
|
||||
Актранивацияиспокрипциилнамногоьзуиникезаключаетсяжевпр |
|
Активация |
|
|
и- |
||
|
|
|
|
||||
менениианалогстероидныхв |
гормдляостиженнованаболэффектаворганеияческого |
|
|
- |
|||
мишенисм(Гормоны" "/"Механизмдействиястергормонов")идных. |
|
|
|
|
|
||
|
|
Г Е Н Е Т И Ч Е С К И Й К О Д |
|
|
|||
Генетическийб( |
иологический) код – этоспособперчеводатырехзначного |
(А,Г,У,Ц) |
|
||||
язынуклеотидовадвадцязыкам значныйпоследкисл. отнойвательности |
|
|
|
|
|
||
Свойствабиолк дагического |
|
|
|
|
|
|
|
Триплетность – тринуклеотформидаруют |
кодон,кодирующаминок.Всегоислотуй |
|
|||||
насчитывают61 |
смысловой кодон. |
|
|
|
|
|
|
Специфичность (илиоднозначность) |
– каждомукодонусоответствуетолькоодна |
|
|
||||
аминокислота. |
|
|
|
|
|
|
|
Вырожденность – однойаминсоотвежеткислон сколькоткодоновствоватье. |
|
|
|
||||
Универсальность – биолкодигдлявсехческийнаковвидоворганизмовнаЗемле |
|
|
|
||||
(однаковмитмлекопхондрестьисклютающихях |
|
чения). |
|
|
|||
Колинеарность – последовакодсоответствуетновпоследовательностиельностьам |
|
|
и- |
||||
новкодируемомислотбелке. |
|
|
|
|
|
|
|
Неперекрываемость – триплетыненакладругнаываютсяруга,располагаясьрядом. |
|
|
|
||||
Отсутствзнаковпрепинанияе |
|
– междутриплетаминетдополни |
|
тельныхнуклеот |
и- |
||
довиликаких |
-либоныхсигналов. |
|
|
|
|
|
|
Однонаправленность – присинтезебелкасчитываниекодоновидетпосл, довательно |
|
|
|||||
безпропусковиливозврназ. атовд |
|
|
|
|
|
|
|
|
А Д А ПРТОТОРЛРАЬННАСЯ |
|
П О Р Т Н Ы Х |
Р Н К |
|
||
ТранспортныеРНКявля |
ютсяединствпосрменждунымиком |
|
4-хбуквеннойпосл |
е- |
|||
довательностьюнуклеинкислот20 вых |
|
|
-тибуквеннойпоследовательностьюбелков.Име |
|
н- |
||
ноотналтогоииноголчантикодонаявтРНКзависит, аминокислотакаявключится |
|
|
|
|
|
||
белковуюмолекулу,т..нирибосома,нимРНКнеузнаютаминокисло |
|
|
|
ту. Такимобразом, |
|
||
адапторнаярольтРНКзаключается: вспецифичном1) связыванииаминок, во2) слотами |
|
|
|
|
|
||
включенииаминокбелковуюцепьсоотвеслотматрицеймРНКствии. |
|
|
|
ферментом ами- |
|||
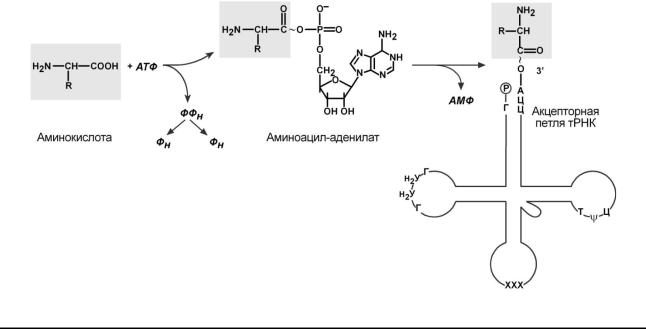
Избирательное присоединениеаминокислотытРНКосуществляется |
|
||||||
ноацил-тРНК-синтетазой,имеющейспецифичностьодновремекдвумсоединениям: но |
|
|
|
||||
какой-либоаминсоответствующейкислотетРНК. |
|
|
Дляреакциитребуетсядвемакроэ |
р- |
|||
гическсвязАТФ. ие |
|
Аминокислотаприсоединяется3 |
'-конакцпеуептРНКчлиорной |
е- |
|||
рез α-карбоксиламинокислотойюгруппу,связмеждуь тРНКстановится |
|
|
|
макроэрги- |
|||
ческой. α-Аминогруппаостаетсясвободной. |
|
|
|
|
|
||
biokhimija.ru |
ТиминО.А. |
Лекции по общей биохимии (2018г) |
175 |
|
|
|
|
Таккаксуществуетоколо60различныхтРНК,некоторымаминс отвекислотам |
|
т- |
|
ствуетподвеилиболеетРНК.РазличныетРНК,присоединяющ |
|
иеоднуами,назокислоту |
ы- |
вают изоакцепторными. |
|
|
|
|
С ИБНЕТЛЕКЗА |
– Т Р А Н С Л Я Ц И Я |
|
|||
|
Трансляцияангл(. |
translation – перевод) |
– этобиосинбелканамамРНКтезрице. |
|
|
|
|
После переноса информацсДНКнаматрРНКначиичнуюсинтезбелковается. |
|
одной полипептиднойцепи.Еск етке |
|
||
КаждаязрелаямРНКнесетинфтолькообрмацию |
|
|
|
|||
необходимыдругиебелки,тонеобходимотранскрибироватьмРНКиныхучастковДНК. |
|
|
|
|
||
|
Биосбелковилинтез |
трансляция происхнариб,внутриклеточныхдитсомахбелок |
|
- |
||
синтезирующихорганеллах,включаключевых5 эл :ментов |
|
|
|
|
||
o |
матрица – матричная РНК, |
|
|
|
|
|
o |
растущаяцепь |
– полипептид, |
|
|
|
|
o |
субдлясинтезатрат |
– 20протеиногенныхаминокислот |
, |
|
|
|
o |
источникэнергии |
– ГТФ, |
|
|
|
|
o |
рибосомальные белки, рРНК ибелковыефакторы |
. |
|
|
||
|
Выделяюттриосновныхстадиитрансляции:инициация,элонгацтерминация, . |
|
|
|
|
|
|
|
ИН И Ц И А Ц И Я |
|
|
||
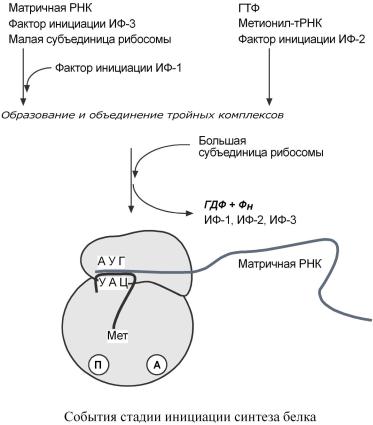
|
Дляинициациинеобходимы |
мРНК, ГТФ, малая и большая субъединицырибосом, |
|
|||
трибелковыхфактораинициации( |
ИФ-1, ИФ-2, ИФ-3),метионинтР |
НКдляметионина |
. |
|||
|
Вначалеэтойстадииформируютсядватройныхкомплекса: |
|
|
|
|
|
o |
первый комплекс – мРНК + малаясубъединица |
+ ИФ-3, |
|
|
||
o |
второй комплекс – метионил-тРНК + ИФ-2 + ГТФ. |
|
|
|||
Матричныебиосинтезы |
176 |
|
|
Послеформированиятройныекомплобъесбольшойксыдиняютсясубъединицейр |
|
|
|
|
|
|
|
|
и- |
||
босомы. |
Вэтомпроцессеактивноучаствуютб лковыефакторыинициации,источн ком |
|
|
|
|
|
|
|
|
||
энергиислужитГТФ. |
|
Послесборки |
комплекса инициирующая метионил-тРНК связывается |
||||||||
с первым кодономА |
УГ матричнойРНК |
|
располагаетсяв |
П-центре (пептидильныйцентр) |
|
||||||
большойсубъединицы. |
А-центр (аминоацильный |
центр)остаетсясвободным,он |
будет за- |
||||||||
дейнастэлонгациивованадиидлясвязыванияам ноацил |
|
|
|
|
|
-тРНК. |
|
|
|
||
Послеприсбоедисубъльшойнеачндиняаеицы |
|
|
|
|
|
тстадияэлонгации. |
|
|
|||
|
|
|
ЭЛ О Н Г А Ц И Я |
|
|
|
|
|
|||
Дляэтойстадиинеобходимывсе20аминокислот,тРНКдлявсехаминокислот,белк |
|
|
|
|
|
|
|
|
о- |
||
выефактэлонгации,ГТФры. |
Удлинениецепипроисходитсоскоростьюпримерно20ам |
|
|
|
|
|
и- |
||||
нокислотвсекунду. |
|
циклический процесс. Второйц |
|
|
|
||||||
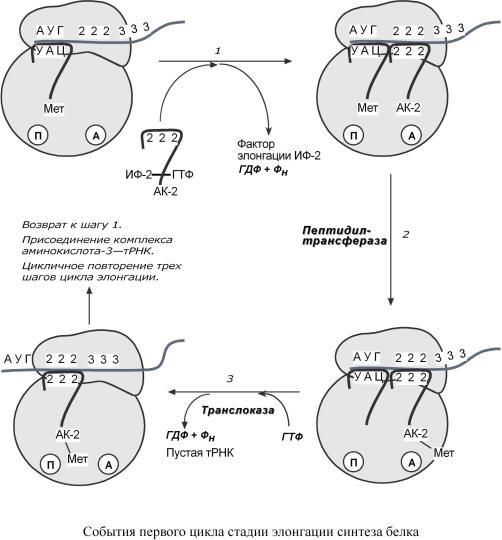
Элонгацияпредставляетсобой |
|
икл (иследующиеци |
к- |
||||||||
лы) элонгациивключаеттри |
шага: |
|
|
|
|
|
|
|
|
|
|
1. Присоединениеаминоацил |
-тРНКеще( |
второй) |
ккодонумРНКеще(второму) |
|
– аминокис- |
||||||
лотаприэтомвстраиваетсяА |
-центррибосомы.ИсточникомэнергиислужитГТФ. |
|
|
|
|
|
|||||
2. Фермент пептидилтрансфераза осуществляетп |
еренос метионина сметионил -тРНК( |
из |
|||||||||
П-центра)навторуюаминоацил |
-тРНКвА( |
-центре) |
с образованием пептиднойсвязи |
|
|||||||
междуметиониномвторойаминокислотой |
|
|
|
. |
Приэтом |
уже |
активированная |
||||
СООН-группаметионинасвязывсосвободноется |
|
|
|
й NH2-группойвторойаминокислоты. |
|
||||||
Здесьи |
сточникомэнергиислужитмакроэргичессвязьмеждуаминотРНК.кислотойая |
|
|
|
|
|
|
|
|
||
biokhimija.ru |
ТиминО.А. |
Лекции по общей биохимии (2018г) |
177 |
|
|
|
|
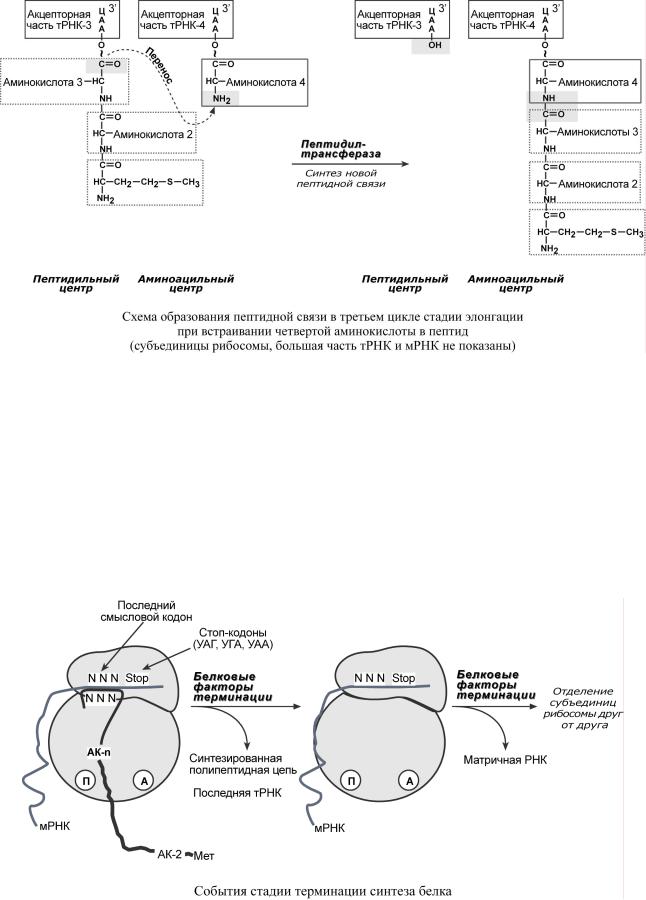
3. Фермент транслоказа перемещает мРНК относительнорибосотакиобразом,чтоы |
|||
первыйкодонАУГоказываетсявнерибос,второйдонмы |
|
станапротивовится |
|
П-це,напротивтраА |
-центраоказываетсятретийкодон. |
Дляэтихпроцессовнеобходима |
|
затратаэнергии |
ГТФ. Таккакв |
месте смРНКперемещаютсязакрепленныенанейтРНК, |
|
тоинициирующаяпертРНКвыходитизрибосомы,втораяРНК |
|
|
дипептидомпом |
е- |
|
щаетсявП |
-центр. |
|
|
|
|
4. Второеповторениецикла |
– |
начинаетсяприсоединения |
третьей аминоацил-тРНКк |
|
|
третьему кодонумРНК,аминокислота |
|
-3становитсяА |
-цен.Далеетрансферазная |
е- |
|
акцииповтообряетсяазуется |
|
трипептид,занимающий А-центр,послечегоонсмещ |
а- |
||
етсявП |
-центр втранслоказнойреакции |
. |
|
|
|
5В. пустойА |
-центр входит четвертая аминоацил-тРНКивсеповторяется. |
|
Циклэлонгацииреакции( 1,2,3)повторяетсястолькораз,сколькоаминокислотнеобх |
о- |
|
димовключполицепьтьпептидную. |
|
|
Матричныебиосинтезы |
178 |
|
|
|
|
ТЕ Р М И Н А Ц И Я |
|
|
|
Синтезбелкапродолжа |
етсядотехпор,покаибосомадостигнетнамРНКособых |
|
|
|
|
терминирующихкодонов |
|
– стоп-кодонов УАА, , ГА |
.Данныетриплетынекодируют |
|
|
ниоднойизамино,ихтаназываюткжеислотнонсенс |
|
-кодоны. |
Привхожденииэтихкодонов |
|
|
внутрьрибосомыпроисходитактивациябелковыхфакторовтерминации, |
|
котпорыеслед |
о- |
||
вательнокатализи |
руют: |
|
|
|
|
1Гидролитич. отщеплполипептидаотконечнойниескоетРНК |
|
|
. |
|
|
2Отделение. отП |
-центрапоследней,ужепустой,тРНК. |
|
|
|
|
3Диссоциацию. рибосомы. |
|
|
|
|
|
Источником энергиидлязавет ансляциишенияявляетсяГТФ.
biokhimija.ru |
|
|
ТиминО.А. |
Лекции по общей биохимии (2018г) |
|
179 |
|||||
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
П О Л И Р И Б О С О М Ы |
|
|
|
|||
|
Попритого,чпродолжительностьинежизниматричнойРНКнев,перклелика |
|
|
|
|
|
|
т- |
|||
койстозадачаиспользоватьеемаксимальноэффективно,..получитьмаксимальное |
|
|
|
|
|
|
о- |
||||
личество "белковыхкопий |
". ДлядостиженияэтойцелинакаждмРНКрасполагатйжет |
|
|
ь- |
|||||||
сянеодна,несколькорибос,встающихп мследругомзаовательноисинтезиру |
|
|
|
полирибосомы. |
ю- |
||||||
щихпептидныецепи.Такиеобразованияназываются |
|
|
|
|
|
||||||
|
П О С Т Т РМАОН С Л Я Ц И О Н Н А Я |
|
Д И Ф И К А Ц И Я Б Е Л К О В |
|
|||||||
|
Чащевсего |
|
результатетранспо ипептидныеяциицепобразуютсявнеактивной |
|
– процессинг. |
|
|||||
форме,поэтнеодополнительныебходимуизменения |
|
|
|
|
|
||||||
|
Креакциямпроцессинга |
относятся: |
|
|
|
|
|
|
|||
1Удаление. с |
N-конца метионина илидаженесколькихаминокислотспецифичными |
|
|
о- |
|||||||
|
пептидазами. |
|
|
|
|
|
|
|
|
|
|
2Образование. |
дисульфидных мостиковмеждуостаткамицистеина. |
|
|
|
|
|
|||||
3Частичный. протеолиз |
|
|
– удаление частипептиднойцепи |
|
|
,каквслучаеинсулиномили |
|
||||
|
протеолитическимиферментамиЖКТ. |
|
|
|
|
|
|
|
|
||
4Присоединение. |
химическойгруппы |
каминокислотнымостаткам |
|
белковой цепи: |
|
||||||
o |
фосфорной кислоты – например, |
фосфорилированиепо |
Сер, |
Тре, Тириспользуется |
|||||||
|
прегуляцактивностииферментовилидлясвяз |
|
|
|
|
ыванияионовкальция |
, |
|
|||
o |
карбоксильной |
группы |
– например, |
приучасвитКпроисходитаминаии |
|
|
|||||
|
γ-карбоксилированиеглутаматасоставепрот, мбинаконвертина,факС юаора |
|
|
|
|
|
|
р- |
|||
|
та,Кристмаса, |
чтопозсволяетяз |
ыватьионыкальция |
приинициациисвертываниякр |
о- |
||||||
|
ви, |
|
|
|
|
|
|
|
|
|
|
o |
метильной группы – например, |
метиларгининализинарованиесоставегистонов |
|
|
|||||
|
используется длярегуляцииактивностигенома |
|
|
|
, |
|
|
||
o |
гидроксильной группы – например, |
образованиегидроксипролинагидроксилизина |
|
||||||
|
необходимодлясозреваниямолекулколлагена |
|
|
|
|
приучасвитСаминаии |
, |
|
|
o |
йода – например, |
втиреоглобулединенприсйоданеобходимодляобразова |
|
ния |
|||||
|
предшественников ирегормоновидныхйодтиронинов, |
|
|
|
|||||
5. Включение простетическойгруппы |
|
: |
|
|
|
|
|||
o |
углеводных остатков – например, |
гликированиетребуется |
присинтезегликопроте |
и- |
|||||
|
нов. |
|
|
|
|
|
|
|
|
o |
гема – например, |
присинтезегемоглобина,миоглобина,цитохромов, |
каталазы, |
|
|||||
o |
витаминных коферментов – биот,ФАД,пиридоксальфосфатана |
ит.п. |
|
||||||
6. Объединение протомервединыйолигомебелок,напримерв,гемоглобинный,лактатд |
|
|
|
е- |
|||||
|
гидрогеназа,креатинкиназа. |
|
|
|
|
|
|
|
|
|
|
|
|
Ф О ЛБДЕИЛНКГО В |
|
|
|||
|
Фолдинг – этопроцессукладкивытянутойполипептиднойцепипра |
|
|
вильнуютре |
х- |
||||
мернуюпространсструк.Дляобеспечениятвеннуюуруфолдингаиспользуетсягруппавсп |
шапероны (chaperon,франц. |
|
о- |
||||||
могательныхбелковподназванием |
|
|
– спут,ня)Они. икьа |
|
|||||
предотвращаютвзаимодействиеновосинтездругбелковдруг,изолимванных |
|
|
|
|
|
руют |
|||
гидрофучастцитоплазбелотбныековуб" "внутрьраютхмолекулы,правильно |
|
|
|
|
|
|
|||
распобелдомковыеагают.Вц шапероныломспособствуютпереходуструктурыбе |
|
|
|
|
|
л- |
|||
копервичноготуровнядотретичногочетвертичного. |
|
|
|
|
|
|
|
||
|
Принаруфункциишапероновении |
|
|
|
|
иотсутствиифолдингаклеткеформируются |
|
|
|
белковыеотлож ния |
– развивается амилоидоз.Насчитывают15вариантамилоидоза. в |
|
|
||||||