

Окислительное фосфорилирование адф. Механизм сопряжения окисления и фосфорилирования. Коэффициент окислительного фосфорилирования (р/о).
Так как электроны всегда стремятся переходить от электроотрицательных систем к электроположительным, их транспорт по ЦПЭ к кислороду сопровождается относительно большим снижением свободной энергии.
В ЦПЭ можно выделить 3 участка, в которых перенос электронов сопровождается относительно большим снижением свободной энергии. Это количество свободной энергии необходимо для синтеза АТФ из АДФ и фосфата (фосфорилирование).
Процесс переноса электронов по ЦПЭ и синтез АТФ энергетически сопряжены.
Синтез АТФ из АДФ и Н3РО4 за счет энергии переноса электронов по ЦПЭ называют окислительным фосфорилированием.
Механизм сопряжения окончательно не выяснен, наиболее обоснованной является хемиосмотическая теория Митчелла, предложенная в 1961г.
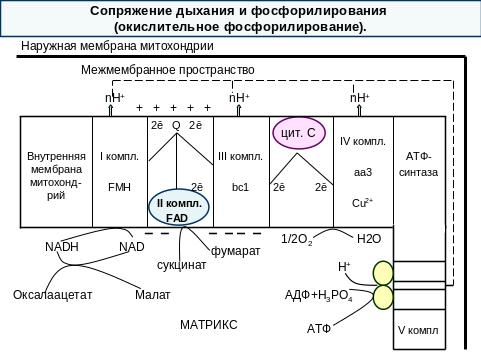
Перенос электронов по ЦПЭ от НАДН к кислороду сопровождается выкачиванием протонов из матрикса митохондрий через внутреннюю мембрану в межмембранное пространство.
Протоны, перенесенные из матрикса в межмембранное пространство, не могут вернуться обратно в матрикс, так как внутренняя мембрана не проницаема для протонов.
Таким образом, создается протонный градиент, при котором концентрация протонов в межмембранном пространстве больше, а рН меньше, чем в матриксе. Кроме того, каждый протон несет положительный заряд, и вследствие этого появляется разность потенциалов по обе стороны мембраны: отрицательный заряд – на внутренней стороне, положительный заряд – на внешней. В совокупности электрический и концентрационный градиенты составляют электрохимический потенциал ΔμН+ - источник энергии для синтеза АТФ.
Энергия электрохимического потенциала (ΔμН+) используется для синтеза АТФ, если протоны возвращаются в матрикс через ионные каналы АТФ-синтазы (V комплекс).
Наиболее активный транспорт протонов в межмембранное пространство, необходимый для образования ΔμН+ происходит на участках ЦПЭ, соответствующих расположению комплексов I, III, IV. Эти участки называют пунктами сопряжения дыхания и фосфорилирования, где и происходит синтез АТФ.
V комплекс – атф-синтаза.
Это интегральный белок внутренней мембраны митохондрий. Он расположен в непосредственной близости к дыхательной цепи. АТФ-синтаза состоит из двух белковых комплексов.
Гидрофобный комплекс F◦ погружен в мембрану. Он служит основанием, которое фиксирует АТФ-синтазу в мембране.
Комплекс F0 состоит из нескольких субъединиц, образующих канал, по которому протоны переносятся в матрикс.
Комплекс F1 выступает в матрикс. Он состоит из 9 субъединиц. Между α- и β- субъединицами располагаются три активных центра, в которых происходит синтез АТФ.
Повышение концентрации протонов в межмембранном пространстве активирует АТФ-синтазу. Электрохимический потенциал ΔμН+ заставляет протоны двигаться по каналу АТФ-синтазы в матрикс. Параллельно под действием ΔμН+ происходят конформационные изменения в парах α- и β- субъединиц белка F1 , в результате чего из АДФ и Н3РО4 образуется АТФ. Электрохимический потенциал, генерируемый в 3 пунктах сопряжения в ЦПЭ, используется для синтеза одной молекулы в каждом пункте.
Отношение количества Н3РО4, использованной на фосфорилирование АДФ, к атому поглощенного кислорода, называют коэффициентом окислительного фосфорилирования и обозначают Р/О.
Для субстратов, которые отдают протоны и электроны на НАД-зависимые дегидрогеназы, коэффициент фосфорилирования равен 3/1=3АТФ, так как протоны и электроны транспортируются через 3 пункта сопряжения (I, III, IV комплексы).
Например, изоцитрат, малат, ПВК и др.
Для субстратов, которые отдают протоны и электроны на ФАД-зависимые дегидрогеназы и сукцинат-фумарат-дегидрогеназу (II комплекс), коэффициент фосфорилирования равен 2/1=2АТФ, так как электроны поступают на III комплекс, минуя первый пункт сопряжения (I комплекс). Например, сукцинат, глицерол-3-фосфат и др.