

Вопрос 27. Судьба ацетил-КоА
(Из лекции) здесь без подробностей. В след вопросах будет освещено:
В ЦТК – при этом образуется 12 молекул АТФ(см пред вопрос).
Жирные кислоты - Ацетил-КоА в цитоплазме служит исходным субстратом для их синтеза
Синтез кетоновых тел
Биосинтез холестерина.
Вопрос 28. Локализация ферментов -окисления жирных кислот. Транспорт жирных кислот в митохондрии. Карнитин-ацилтрансфераза.
Как уже было сказано, биосинтез жирных кислот протекает в цитозоле, а окисление - в митохондриях.
Ферменты:
1)Ацил-КоА синтетазы (необходимы для активации жирных кислот) находятся как в цитозоле, так и в матриксе митохондрий. Эти ферменты отличаются по специфичности к жирным кислотам с различной длиной углеводородной цепи. Жирные кислоты с короткой и средней длиной цепи (от 4 до 12 атомов углерода) могут проникать в матрикс митохондрий путём диффузии. Активация этих жирных кислот происходит в матриксе митохондрий. Жирные кислоты с длинной цепью, которые преобладают в организме человека (от 12 до 20 атомов углерода), активируются ацил-КоА синтетазами, расположенными на внешней мембране митохондрий.
Транспорт жирных кислот с длинной углеводородной цепью в митохондриях
β-Окисление жирных кислот, происходит в матриксе митохондрий, поэтому после активации жирные кислоты должны транспортироваться внутрь митохондрий. Жирные кислоты с длинной углеводородной цепью переносятся через плотную внутреннюю мембрану митохондрий с помощью карнитина. Карнитин поступает с пищей или синтезируется из незаменимых аминокислот лизина и метионина. В реакциях синтеза карнитина участвует витамин С (аскорбиновая кислота).
В наружной мембране митохондрий находится фермент карнитинацилтрансфераза I (карнитин-пальмитоилтрансфераза I), катализирующий реакцию с образованием ацилкарнитина.
Образовавшийся ацилкарнитин проходит через межмембранное пространство к наружной стороне внутренней мембраны и транспортируется с помощью карнитинацилкарнитинтранс-локазы на внутреннюю поверхность внутренней мембраны митохондрий, где фермент карнитинацилтрансфераза II катализирует перенос ацила на внутримитохондриальный КоА (рис. 8-26). Таким образом, ацил-КоА становится доступным для ферментов β-окисления. Свободный карнитин возвращается на цитозольную сторону внутренней мембраны митохондрий той же транслоказой.
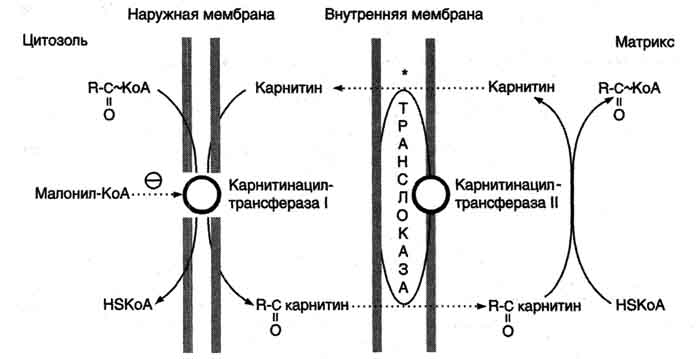
Рис. 8-26. Перенос жирных кислот с длинным углеводородным радикалом через мембраны митохондрий. Фермент карнитинацилтрансфераза I - регуляторный фермент β-окисления; ингибируется малонил-КоА - промежуточным метаболитом, образующимся при биосинтезе жирных кислот. * - карнитинацилкарнитинтранслоказа.
Теперь по формулам:
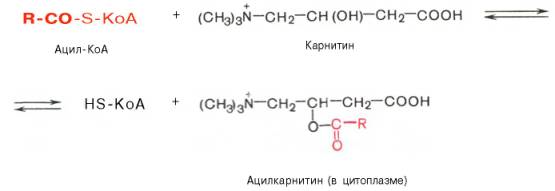
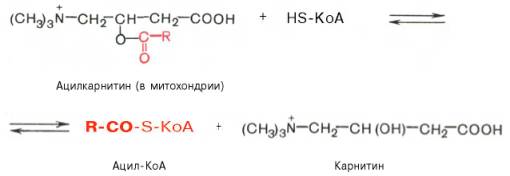
На внутренней поверхности внутренней мембраны находится фермент карнитинацил трансфераза II, катализирующий обратный перенос ацила с карнитина на внутримитохондриальный КоА. После этого ацил-КоА включается в реакции β-окисления.
Ну, логично предположить, что все остальные ферменты, участвующие в В-окислении, находятся в матриксе митохондрий, так как сам процесс окисления и проходит в матриксе.