

Глава 10
ВОЛОКНИСТЫЕ СОЕДИНИТЕЛЬНЫЕ ТКАНИ
ОБЩИЕ СВЕДЕНИЯ
Волокнистые соединительные ткани являются наиболее типичными представителями группы соединительных тканей, отчего их называют также собственно соединительными тканями. Как и другие ткани этой группы, они характеризуются высоким содержанием межклеточного вещества. В последнем значительное место занимают волокна (что отражено в наименовании этих тканей), которые выполняют важную функциональную роль; пространства между волокнами заполнены основным аморфным веществом. Межклеточное вещество продуцируется клетками волокнистых соединительных тканей.
Функции волокнистых соединительных тканей включают все основные функции, свойственные соединительным тканям (см. главу 6), однако наиболее важными из них являются: (1) трофическая, (2) регуляторная, (3) защитная и (4) опорная (механическая). Биологические и физико-химические свойства, определяющие функции конкретного вида волокнистых соединительных тканей, отражены в характеристиках, которые положены в основу их классификации.
КЛАССИФИКАЦИЯ
ВОЛОКНИСТЫХ СОЕДИНИТЕЛЬНЫХ
ТКАНЕЙ
Классификация волокнистых соединительных тканей основана на соотношении клеток и межклеточного вещества, а также свойствах и особенностях организации (степени упорядоченности) последнего (см. главу 6). В соответствии с классификацией выделяют рыхлую волокнистую соединительную ткань и плотную волокнистую соединительную ткань.
1. Рыхлая волокнистая соединительная ткань характеризуется сравнительно невысоким содержанием волокон в межклеточном веществе, относительно большим объемом основного аморфного вещества, многочисленным и разнообразным клеточным составом.
2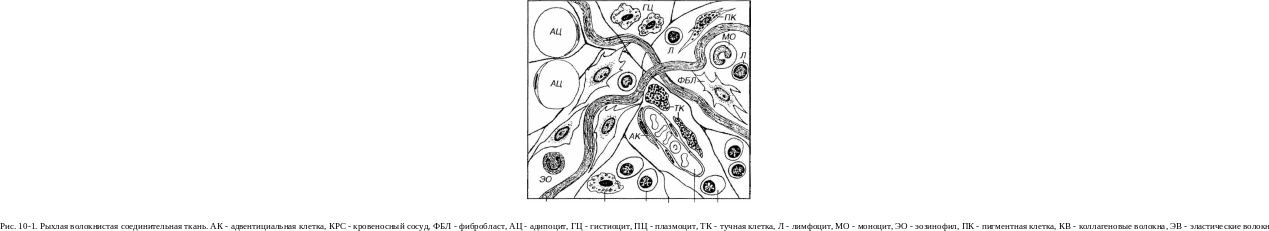 .
Плотная
волокнистая соединительная ткань
отличается
преобладанием в межклеточном веществе
волокон при незначительном объеме,
занимаемом основным аморфным веществом,
относительно малочисленным и
однообразным клеточным составом.
Плотную волокнистую соединительную
ткань, в свою очередь, подразделяют на:
.
Плотная
волокнистая соединительная ткань
отличается
преобладанием в межклеточном веществе
волокон при незначительном объеме,
занимаемом основным аморфным веществом,
относительно малочисленным и
однообразным клеточным составом.
Плотную волокнистую соединительную
ткань, в свою очередь, подразделяют на:
(а) оформленную (в которой все волокна ориентированы в одном направлении) и
(б) неоформленную (с различной ориентацией волокон).
Так как в рыхлой волокнистой соединительной ткани волокна всегда имеют разнообразный ход, она является неоформленной, однако обычно это не отмечается в ее названии, поскольку оформленного варианта этой ткани не существует.
РЫХЛАЯ ВОЛОКНИСТАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ
Рыхлая волокнистая соединительная ткань является самым распространенным видом соединительных тканей и имеет наиболее типичное для этих тканей строение, так как содержит разнообразные клетки и все компоненты межклеточного вещества (рис. 10-1). Она выполняет все функции, свойственные соединительным тканям, взаимодействуя с другими тканями, связывая их между собой (что оправдывает общее название этой группы тканей) и способствуя поддержанию гомеостаза в организме. Эта ткань обнаруживается повсеместно, во всех органах - она образует их строму (основу), в частности, междоль-ковые прослойки и прослойки между слоями и оболочками, заполняет пространства между функциональными элементами других тканей, сопровождает нервы и сосуды, входит в состав кожи и слизистых оболочек.
КЛЕТКИ РЫХЛОЙ ВОЛОКНИСТОЙ СОЕДИНИТЕЛЬНОЙ ТКАНИ
Клетки рыхлой волокнистой соединительной ткани представляют собой сложную гетерогенную популяцию функционально разнообразных и взаимодействующих между собою и с компонентами межклеточного вещества элементов (см. рис. 10-1), которые условно объединяют в несколько групп.
КВ ГЦ ПЦ ЭВ КРС пц
По признаку постоянства присутствия в составе рыхлой волокнистой соединительной ткани ее клетки подразделяют на:
оседлые (фиксированные, резидентные) клетки, т.е. образующиеся и постоянно пребывающие в этой ткани. К этой группе относят адвентициалъные клетки, фибробласты, фиброциты и жировые клетки (адипоциты). В зрелой рыхлой волокнистой соединительной ткани содержание оседлых клеток относительно стабильно;
блуждающие клетки (иммигранты) - подвижные элементы, поступающие в соединительную ткань из крови. В эту группу включают все виды лейкоцитов (гранулоцитов и агранулоцитов). Содержание этих клеток в отдельных участках соединительной ткани может существенно изменяться при различных иммунных реакциях и воспалении.
Макрофаги (гистиоциты), плазматические и зучггые клетки одни авторы считают оседлыми элементами (поскольку они образуются в соединительной ткани и постоянно присутствуют в ней), другие причисляют к блуждающим клеткам (так как они дифференцируются из предшественников, циркулирующих в крови).
По источникам развития выделяют три группы клеток:
Клетки линии механоцитов - адвентициальные клетки, фиб-робласты, фиброциты, адипоциты - развиваются из особой стволовой клетки этой клеточной линии, которая имеет мезенхимное происхождение. К линии механоцитов помимо указанных клеток рыхлой волокнистой соединительной ткани относят клетки других тканей - ретикулярной (ретикулярные клетки), а также скелетных соединительных (хондроциты и остеоциты), поскольку вырабатываемые ими продукты (компоненты межклеточного вещества) обеспечивают механические свойства тканей.
Клетки-потомки стволовой клетки крови (СКК) - макрофаги (гистиоциты), дендритные АПК, плазматические и тучные клетки, лейкоциты (гранулоциты и агранулоциты) - развиваются, как следует из названия группы, из СКК, которая происходит из мезенхимы.
Клетки нейралъного происхождения - ггягментные клетки (развиваются из предшественников, которые выселяются из нервного гребня).
Фибробласты
Фибробласты (от лат. fibra - волокно и греч. blastos - росток) -наиболее распространенные и функционально ведущие клетки рыхлой волокнистой соединительной ткани, относящиеся к клеточной линии механоцитов.
Функции фибробластов (лишь частично отражены в их названии):
Ф Продукция всех компонентов межклеточного вещества (волокон и основного аморфного вещества);
0 Поддержание структурной организации и химического гомео-стаза межклеточного вещества (за счет сбалансированных процессов его выработки и разрушения);
G) Регуляция деятельности других клеток соединительных тканей и влияние на другие ткани.
Развитие фибробластов
Источником развития фибробластов в эмбриогенезе является мезенхима. После рождения фибробласты представляют собой сложную систему (дифферон) клеток, имеющих общего предшественника и различающихся по степени дифференцировки, морфологическим и функциональным характеристикам. Основная линия развития в этом диффе-роне (рис. 10-2) представлена последовательностью:
с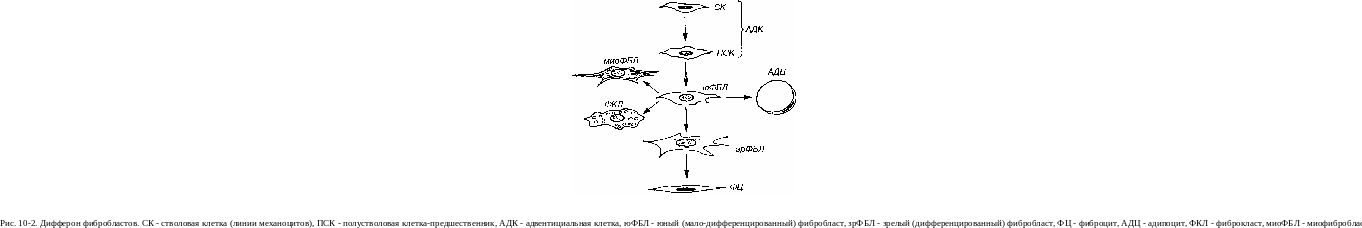 тволовая
клетка линии механоцитов -> полустволовая
клетка-предшественник - >
малодифференцированный (юный) фибробласт
-> зрелый (дифференцированный) фибробласт
-> фиброцит.
тволовая
клетка линии механоцитов -> полустволовая
клетка-предшественник - >
малодифференцированный (юный) фибробласт
-> зрелый (дифференцированный) фибробласт
-> фиброцит.
Стволовая клетка линии механоцитов и полустволовые клетки-предшественники, образующиеся из нее в ходе дифференци-ровки, представляют собой наиболее ранние элементы дифферона фиб-робластов. Морфологически им, по всей видимости, соответствует ад-вентициалъная клетка - мелкая веретеновидная уплощенная мало-дифференцированная клетка, располагающаяся по ходу капилляров (см. рис. 10-1). Для нее характерно темное ядро и базофильная цитоплазма, содержащая слабо развитые органеллы. Стволовые клетки устойчивы к повреждающим воздействиям, редко делятся и образуют самоподдерживающуюся популяцию. Полустволовые клетки при стимуляции способны к высокой митотической активности, однако их синтетический аппарат не развит и они не продуцируют компонентов межклеточного вещества соединительной ткани. Вопрос о природе и свойствах стволовой клетки линии механоцитов окончательно не разработан.
Альтернативные представления о природе стволовых клеток линии механоцитов поддерживаются некоторыми авторами, которые отождествляют их с перицитами - особыми клетками, лежащими кнаружи от эндотелиоцитов в сосудах микропиркуляторного русла. Для перицитов, в отличие от адвентициальных клеток, характерна сложная форма (варьирующая в различных сосудах), наличие первичных (крупных) отростков, отходящих от клеточного тела и разделяющихся на ряд вторичных (более мелких), которые охватывают эндотелиоциты снаружи. Функция перицитов до конца не выяснена; помимо представления о них, как о малодифференцированных клетках линии механоцитов, разные авторы приписывают им выполнение транспортной, сократительной, фагоцитарной и регуляторной функций, способность превращаться в гладкие миоциты и макрофаги, контролировать образование и рост сосудов (ангиогенез).
Малодифференцированный (юный) фибробласт - базофильная клетка более крупных размеров, чем адвентициалъная, с небольшим числом отростков. Для нее характерно крупное круглое или овальное ядро с 1-2 ядрышками, умеренно развитый синтетический аппарат. Она сохраняет способность к пролиферации, но они уже начинает осуществлять синтез типичных компонентов межклеточного вещества соединительной ткани - коллагена и гликозаминогликанов.
Способность юных фибробластов к направленной миграции определяет их важную роль в репаративных процессах, в частности, в заживлении ран. Миграция осуществляется благодаря наличию в их цитоплазме сократимых микрофиламентов, на которые опосредованно передаются сигналы с многочисленных рецепторов плазмолеммы, воспринимаюших молекулы хемотаксических веществ. Факторами, привлекающими их в очаг повреждения, служат продукты, выделяемые макрофагами, Т-лимфоцитами, тромбоцитами (в частности, ТРФР, называемый "раневым гормоном"), фибронектин, а также пептиды, образующиеся при расщеплении коллагена. Многие из этих факторов оказывают на юные фибробласты также митогенное действие, стимулируют их функциональную активность и дифференцировку, по завершении которой эти клетки превращаются в зрелые фибробласты.

Рис. 10-3. Ультраструктурная организация фибробласта (ФБЛ) и фиброцита (ФЦ). ЩС - щелевое соединение (между отростками ФБЛ и ФЦ), КГ - комплекс Голь-
джи.
Зрелый (дифференцированный) фибробласт - крупная (на пленочных препаратах - более 40-50 мкм в поперечнике) отростчатая клетка с нерезкими границами и светлым ядром, содержащим мелкодисперсный хроматин и 1-2 ядрышка (см. рис. 10-1 и 10-3). Цитоплазма слабо базофильна и характеризуется диплазматической дифференци-ровкой - нерезким разделением на внутреннюю, более плотную часть, окружающую ядро, - эндоплазму и периферическую, сравнительно светлую и образующую отростки - эктоплазму. Эндоплазма содержит большую часть органелл мощно развитого синтетического аппарата, а эктоплазма заполнена преимущественно элементами цитоскелета. Цистерны грЭПС часто растянуты, содержат мелкозернистый материал низкой электронной плотности. В цитоплазме располагаются также лизосомы, митохондрии, липидные капли и многочисленные пузырьки. Все элементы цитоскелета хорошо выражены. Фибробласт обладает подвижностью, способностью изменять свою форму и обратимо прикрепляться к другим клеткам и компонентам межклеточного вещества (волокнам).
Функции зрелого фибробласта заключаются в сбалансированных процессах продукции, перестройки и частичного разрушения межклеточного вещества (см. ниже), что обеспечивает возможность тонкой регуляции его архитектоники и состояния. Фибробласты оказыват также влияние на деятельность клеток других типов в соединительной и соседних с ней тканях.
Регуляция деятельности фибробластов осуществляется факторами, вырабатываемыми макрофагами, Т-лимфоцитами, тромбоцитами и эпителиальными клетками (включая эндотелиоциты), а также различными гормонами.
Регуляторное влияние фибробластов на другие клетки обеспечивается благодаря продукции ими гуморальных факторов, активно воздействующих на рост, дифференцировку и функциональную активность как их собственной популяции, так и макрофагов, моноцитов, лимфоцитов, гладкомышечных и эпителиальных клеток. На указанные клетки в качестве локальных регуляторов воздействуют также вырабатываемые фибробластами компоненты межклеточного вещества (в особенности, фибронектин, гликозаминогликаны, коллагены различных типов).
Большинство фибробластов разрушается в процессе жизнедеятельности, но часть их превращается в малоактивную долгоживущую форму - фиброциты.
Фиброцит - конечная форма развития фибробласта - узкая веретенообразная, неспособная к пролиферации клетка с длинными тонкими отростками, которые часто имеют уплощенную крыловидную форму. Ядро - сравнительно плотное (с преобладанием гетерохроматина), занимает большую часть клетки. Цитоплазма содержит слабо развитый синтетический аппарат, значительное количество лизосом, липофусщшо-вых гранул (см. рис. 10-3). Функция этих клеток состоит в регуляции метаболизма и поддержании стабильности межклеточного вещества; синтез его компонентов осуществляется ими очень слабо. Фиброциты располагаются между пучками коллагеновых волокон.
Фибропласты (от лат. fibra - волокно и греч. klasis - разрушение) - клетки дифферона фиброцитов, специализированные на функции разрушения межклеточного вещества соединительной ткани, которая резко преобладает над их синтетической и секреторной активностью. По-видимому, процессы деградации межклеточного вещества этими клетками осуществляются внутриклеточным и внеклеточным механизмами, анало-пгчными тем, что используются зрелыми фибробластами (см. ниже). В их цитоплазме выявляются многочисленные вакуоли, содержащие ли-тические ферменты и коллагеновые фибриллы на различных стадиях лизиса. Эти клетки обеспечивают перестройку и инволюцию соединительной ткани; они особенно многочисленны в молодой соединительной (грануляционной) ткани и рубцах, подвергающихся обратному развитию.
Миофибробласты - особые клетки, которые по своему строению и функции занимают промежуточное положение между типичными фибробластами и клетками гладкой мышечной ткани - гладкими мио-цитами. На светооптическом уровне их невозможно отличить от типичных фибробластов, однако по ультраструктурной организации они близки к гладким миоцитам, хотя, в отличие от последних, и не окружены базальной мембраной. Более половины объема их цитоплазмы занимают элементы сократительного аппарата. Их синтетический аппарат развит слабее, чем в зрелых фибробластах. Иммуноцитохимически в их цитоплазме помимо виментина вьвтвляются актин и десмин гладкомы-шечного типа.
Активация миофибробластов происходит при повреждении соединительной ткани. Они активно участвуют в репаративных процессах: образуют коллаген (главным образом, /// типа), который заполняет и связывает поврежденные участки; сокращаясь, они стягивают края раны и уменьшают ее размеры (контракция раны - от лат. contracüo -сокращение). В связи с указанной функцией миофибробласты в большом количестве обнаруживаются в молодой регенерирующей соединительной (грануляционной) ткани, рубцах, в мышечной оболочке матки при беременности. В ходе заживления раны миофибробласты с высоким содержанием актина постепенно погибают механизмом апоптоза; в рубцах они замещаются типичными фибробластами и фиброцитами. С повышенной активностью миофибробластов связывают развитие ряда заболеваний (фиброза легкого, печени, почек).
Жировые клетки
Жировые клетки (адипоциты), согласно принятым представлениям, образуются из малодифференцированных (юных) фибробластов
(см. рис. 10-2) путем накопления в их цитоплазме мелких липидных капель, которые сливаются между собой в одну крупную, заполняющую ее почти целиком (см. рис. 11-1). Подробное описание развития, строения и функции этих клеток приведено в главе 11.
Жировые клетки являются нормальным компонентом рыхлой волокнистой соединительной ткани и в небольшом количестве встречаются в ней повсеместно, располагаясь поотдельности или в виде мелких скоплений. Ткань, в которой адипоциты являются структурно и' функционально ведущими клеточными элементами, называют жировой и относят к одному из видов соединительных тканей со специальными свойствами (см. главы 6 и 11).
Морфологические признаки гистиоцитов зависят от степени их функциональной активности. В целом, вследствие наличия переходных форм популяция гистиоцитов характеризуются выраженным полиморфизмом.
Покоящиеся гистиоциты трудно идентифицировать на светооп-гическом уровне. Они имеют вид мелких уплощенных клеток удлиненной или отростчатой формы с четкими контурами, прикрепленных к коллагеновым волокнам. Эти клетки характеризуются небольшим темным ядром и плотной цитоплазмой со слабо развитыми органеллами.
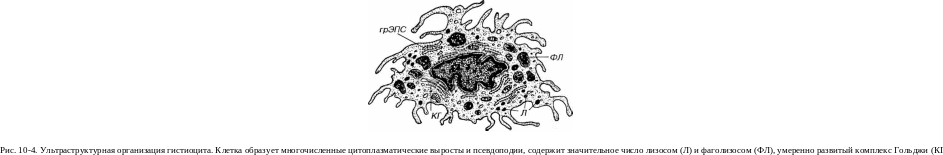
Макрофаги (гистиоциты)
Макрофаги (гистиоциты) - вторые по численности (после фиб-робластов) клетки рыхлой волокнистой соединительной ткани. Они принадлежат к линии потомков стволовой клетки крови и непосредственно образуются из моноцитов после их миграции в соединительную ткань из просвета кровеносных сосудов (см. главу 7). В соединительной ткани макрофаги располагаются поодиночке или фуппами. Эти клетки очень многочисленны в собственной пластинке слизистых оболочек, а также в серозных оболочках. Они могут пребывать в одном из двух взаимообратимых состояний:
(1) покоящихся клеток, обладающих низкой функциональной активностью;
(2) блуждающих клеток с высокой функциональной активностью.
По мнению некоторых авторов, термин гистиоцит следует употреблять только применительно к клеткам в покое, однако в настоящее время он, как правило, используется в более общем смысле для обозначения макрофага соединительной ткани.
Функции гистиоцитов (подробнее см. главу 7):
Ф распознавание, поглощение и переваривание поврежденных, зараженных, опухолевых и погибших клеток, компонентов межклеточного вещества, а также экзогенных материалов и микроорганизмов;
@ участие в индукции иммунных реакций посредством захвата, переработки (процессинга) антигенов и представления их лимфоцитам (играют роль антиген-представляющих клеток);
регуляция деятельности клеток других типов (фибробластов, лимфоцитов, тучных клеток, эндотелиоцитов и др.).
Блуждающие (активные) гистиоциты обладают высокой подвижностью, изменчивой (отростчатой, реже округлой) формой с неровными, но обычно четко выявляемыми краями (см. рис. 10-1). Их ядро светлее, чем в покоящихся клетках, но темнее, чем в фибробластах; в нем может выявляться ядрышко. Цитоплазма содержит многочисленные лизосомы (рис. 10-4) и развитые элементы цитосклета, которые концентрируются в области псевдоподий; другие органеллы развиты умеренно. Многочисленные крупные фаголизосомы, содержащие перевариваемые продукты, в виде вакуолей хорошо видны под световым микроскопом, придавая цитоплазме гистиоцитов вспененный вид. На плазмолемме в большом количестве находятся рецепторы цитокинов, гормонов, хемоаттрактантов, а также адгезивные молекулы, которые обеспечивают контактные взаимодействия гистиоцитов с другими клетками и компонентами межклеточного вещества.
Преобразования гистиоцитов в рыхлой волокнистой соединительной ткани. При активации, происходящей под действием микроорганизмов или их продуктов, а также ряда цитокинов (см. главу 7), клетки в покое могут превращаться в блуждающие. Последние, получая стимулирующие сигналы, способны длительно находиться в состоянии высокой активности, однако в конечном итоге погибают механизмом апоптоза и фагоцитируются другими макрофагами. Под воздействием дополнительных сигналов в очаге повреждения они могут также превратиться в особые виды макрофагов - гигантские многоядерные клетки и эпителиоидные клетки (см. главу 7). Утрачивая активность и подвижность и прикрепляясь к коллагеновым волокнам, блуждающие клетки способны возвращаться в состояние покоя.
Дендритные
антиген-представляющие клетки (АПК)
Дендритные АПК являются постоянными клеточными элементами рыхлой волокнистой соединительной ткани, отаосянгимися к потомкам стволовой клетки крови. По всей видимости, они образуются непосредственно из моноцитов крови после их миграции в ткани. Не исключается полностью и возможность их развития из гематогенного предшественника, отличного от моноцитов. Установлено, что дендритные АПК в организме образуют единую систему морфологически и функционально сходных элементов. Общей функциональной особенностью дендритных АПК служит свойственная им высокая активность захвата, процес-синга и представления антигенов лимфоцитам. Морфологическим признаком, характерным для этих клеток, является их отростчатая форма, наличие многочисленных ветвящихся цитоплазматических отростков, которые могут укорачиваться при перемещении клеток.
Дендритные АПК, выявляемые в соединительной ткани, могут относиться к одной из двух популяций клеток: (1) АПК, специализированным на захвате антигенов только в пределах этой ткани (собственно соединительнотканным АПК), и (2) АПК, располагающимся и захватывающим антигены в эпителиях (кожи, слизистых оболочек), которые находятся в процессе миграции через соединительную ткань из эпителия в лимфатические сосуды или из кровеносных сосудов в эпителий. В ходе миграции происходят изменения ряда фенотипических свойств дендритных АПК. Детали строения, функции и распределения этих клеток рассматриваются в главах 7 и 8.
Тучные клетки
Тучные клетки - постоянный клеточный компонент рыхлой волокнистой соединительной ткани, осуществляющий важные регулятор-ные функции. Относятся к потомкам стволовой клетки крови.
Терминология. Тучные клетки получили свое название в связи с первоначальным ошибочным предположением о том, что их многочисленные гранулы содержат запасы питательных веществ. Этим, вероятно, объясняется и другое их название - лаброциты (от греч. labros -жадный и cytos, или kytos - клетка). Тучные клетки именуют также тканевыми базофилами, подчеркивая их сходство с базофильными гра-нулоцитами крови, однако это название неудачно, так как оно создает путаницу между тучными клетками и отличающимися от них базофилами крови после их миграции в соединительную ткань.
Развитие тучных клеток осуществляется в тканях из предшественника, который имеет, как предполагают, костномозговое происхождение. На их дифференцировку и рост влияют ИЛ-3 (продуцируемый Т-лимфоцитами) и факторы клеточного микроокружения (фибробласты, эпителиальные клетки и их продукты). В отличие от базофилов, которые после миграции в ткани живут недолго (от нескольких часов до нескольких суток), тучные клетки, по-видимому, обладают сравнительно большой продолжительностью жизни (от нескольких недель до нескольких месяцев). В течение этого периода под действием соответствующих стимулов тучные клетки, очевидно, способны делиться.
Функции тучных клеток в целом сходны с функциями базофилов, находящихся в тканях (см. главу 7). К ним относятся:
Ф Гомеостатическая, которая осуществляется в физиологических условиях путем медленного выделения небольших количеств биологически активных веществ, способных влиять на различные тканевые функции - в первую очередь, на проницаемость и тонус сосудов и поддержание баланса жидкостей в тканях.
® Защитная и регуляторная, которая обеспечивается путем локального выделения медиаторов воспаления и хемотаксических факторов, обеспечивающих (а) мобилизацию эозинофилов и различных эф-фекторных клеток, участвующих в так называемых реакциях поздней фазы; (б) воздействие на рост и созревание соединительной ткани в зоне воспаления.
<Э Участие в развитии аллергических реакций вследствие наличия высокоаффинных рецепторов к иммуноглобулинам класса Е (IgE) на их плазмолемме и функциональной связи этих рецепторов с секреторным механизмом.
Р аспределение
тучных клеток в
организме.
Тучные
клетки располагаются преимущественно
около мелких сосудов - периваскулярно
(см.
рис. 10-1), что, вероятно, связано с их
регуляторной функцией и влиянием на
проницаемость сосудов. Распределение
тучных клеток в организме неравномерно
- соединительная ткань различных органов
содержит неодинаковое их количество.
Этими клетками особенно богата дерма
- соединительнотканная
часть кожи, где их содержание достигает
10-20 тыс. клеток/мм3.
Они также очень многочисленны в
собственной пластинке слизистых
оболочек пищеварительного тракта,
дыхательной, выделительной и половых
систем, в строме молочной железы и
тимуса. В среднем, в рыхлой волокнистой
соединительной ткани их относительное
содержание составляет 10% от общего числа
клеток.
аспределение
тучных клеток в
организме.
Тучные
клетки располагаются преимущественно
около мелких сосудов - периваскулярно
(см.
рис. 10-1), что, вероятно, связано с их
регуляторной функцией и влиянием на
проницаемость сосудов. Распределение
тучных клеток в организме неравномерно
- соединительная ткань различных органов
содержит неодинаковое их количество.
Этими клетками особенно богата дерма
- соединительнотканная
часть кожи, где их содержание достигает
10-20 тыс. клеток/мм3.
Они также очень многочисленны в
собственной пластинке слизистых
оболочек пищеварительного тракта,
дыхательной, выделительной и половых
систем, в строме молочной железы и
тимуса. В среднем, в рыхлой волокнистой
соединительной ткани их относительное
содержание составляет 10% от общего числа
клеток.
Проявлением регуляторной функции тучных клеток служит нарастание их количества в строме различных органов, функциональная активность которых повышается, например, в щитовидной железе при ее гиперфункции, в лактирующей молочной железе, в матке при беременности и в течение менструального цикла и т.п. Оно увеличено также вблизи и внутри очагов хронического воспаления, в опухолях и по периферии заживающих ран. Механизмами локального нарастания содержания тучных клеток могут служить их миграция, обусловленная хемо-аттрактантами, усиленная дифференцировка из местных предшественников и, возможно, митотическое деление.
В тканях тучные клетки устанавливают многочисленные адгезивные контакты с фибробластами, эндотелиалъными клетками мелких сосудов, коллагеновьгми и нервными волокнами, молекулами фибронекги-на, ламинина и другими компонентами межклеточного вещества. Эти взаимодействия оказывают регуляторные влияния как на состояние самих тучных клеток (способствует их дифференцировке из предшественников, облегчают их миграцию, распластывание, секреторную реакцию), так и на клетки других типов.
Строение тучных клеток. Тучные клетки имеют удлиненную или округлую форму, неровную поверхость с многочисленными тонкими отростками и выростами. Они в 1.5-2 раза крупнее базофилов (диаметр 20-30 мкм).
Ядро тучных клеток - сравнительно небольшое, несегментирован-ное, овальное или округлое, с умеренным содержанием гетерохрома-тина. На светооптическом уровне оно часто прослеживается с трудом, так как маскируется гранулами, содержапгимися в цитоплазме.
Цитоплазма тучных клеток содержит умеренно развитые органел-лы, элементы цитоскелета, липидные капли и гранулы (рис. 10-5).
-296-
Гранулы тучных клеток сходны по строению и составу содержимого с гранулами базофилов, но не идентичны им. Они также окрашиваются метахромагически, но они мельче, чем в базофилах, более многочисленны и обладают более варабельной формой и ультраструктурой (даже в составе одной клетки). Встречаются гранулы с плотным, крупно- или мелкозернистым гомогенным содержимым, с кристаллоидной структурой, с матриксом умеренной плотности, в который погружены более плотные структуры (иногда в форме "пергаментных свитков"). Последний вид гранул особенно характерен для тучных клеток слизистых оболочек. Нередко обнаруживаются гранулы смешанного строения (см. рис. 10-5).
Содержимое гранул тучных клеток: гепарин, гистамин, дофамин, хемотаксические факторы эозинофилов и нейтрофилов, хондроитин-сульфаты, гиалуроновая кислота, гликопротеины и фосфолипиды. В составе основных белков гранул имеются нейтральные протеазы, кислые гидролазы, катепсин С
Функциональная морфология тучных клеток в физиологических условиях. Феномен медленной дегронуляции тучных клеток человека (длящейся сутками), как и аналогичная реакция базо-
- 297 -
Участие тучных клеток в реакциях поздней фазы (длительной иммунной стимуляции). Разнообразные биологически активные вещества, выделенные тучными клетками, привлекают базофилы, эози-нофилы, нейтрофилы, макрофаги, а также другие клетки и облегчают их миграцию из кровеносных сосудов в ткани, усиливая их адгезию к эндотелию. Выселившиеся клетки секретируют ряд собственных медиаторов, которые могут привлекать новые клетки, поддерживая или усугубляя повреждение тканей. Вместе с тем, некоторые из продуцируемых тучными клетками веществ способствуют течению репаративных процессов, в частности, стимулируют выработку межклеточного вещества фибробластами и ангиогенез.
Вещества, угнетающие дегрануляцию тучных клеток, (с ралич-ным механизмом фармакологического действия) нашли широкое клиническое применение в качестве средств профилактики и лечения аллергических заболеваний.
Плазматические клетки (плазмоциты)
Плазматические клетки (плазмоциты) и их предшественники -В-лимфоциты, находящиеся на различных этапах преобразования в плазмоциты - в небольших количествах постоянно содержатся в различных участках рыхлой волокнистой соединительной ткани (см. рис. 10-1). Они особенно многочисленны в соединительной ткани серозных оболочек, собственной пластинки различных слизистых оболочек, а также вокруг концевых отделов и вьшодных протоков экзокринных желез. Эти клетки имеют мелкие размеры, располагаются поодиночке или группами и обладают высокой синтетической и секреторной активностью, вырабатывая и выделяя антитела (иммуноглобулины) и обеспечивая тем самым гуморальный иммунитет. Характерные морфологические и функциональные признаки плазмоцитов описаны в главе 8.
Лейкоциты
Лейкоциты (гранулоциты и агранулоциты) являются нормальными клеточными компонентами рыхлой волокнистой соединительной ткани, в (или через) которую они мигрируют для выполнения своих функций после выхода из кровеносного русла (см. главу 7). Лимфоциты, в отличие от других видов лейкоцитов, способны из соединительной ткани через оттекающую лимфу вновь попадать в кровь (осуществлять рециркуляцию, см. главу 8).
Содержание лейкоцитов в рыхлой волокнистой соединительной ткани в норме незначительно. Выделяя цитокины, эти клетки могут оказывать регуляторное влияние друг на друга, на остальные виды клеток соединительной ткани и на клетки соседних тканей.
Локальное увеличение числа лейкоцитов в рыхлой волокнистой соединительной ткани, образующих в большей или меньшей степени очерченные скопления, выявляется при воспалении (см. ниже). В частности, при остром воспалении в таких скоплениях (инфильтратах) преобладают нейтрофильные гранулоциты, при хроническом - обнаруживаются преимущественно лимфоциты, плазматические клетки, моноциты и образующиеся из них макрофаги.
Пигментные клетки
Пигментные клетки человека имеют нейральное происхождение и являются потомками клеток, выселившихся в эмбриональном периоде из нервного гребня. Цитоплазма этих клеток содержит пигменты меланины (от греч. те1апоБ - черный). Цвет пигментов варьирует от коричнево-черного (эумеланины, от греч. ей - истинный) до желто-коричневого (фео-меланины, от греч. рпео- светлый). Пигментные клетки имеют отростчатую форму и подразделяются на два вида - меланоциты, которые вырабатывают пигмент, и меланофоры, способные лишь накапливать его в цитоплазме (см. главу 11). Пигментные клетки входят в состав рыхлой волокнистой соединительной ткани (см. рис. 10-1), хотя у человека и других млекопитающих они встречаются в ней сравнительно редко.
Повышенное содержание пигментных клеток характерно для соединительнотканной части кожи (дермы) некоторых участков тела (мошонки, сосков, перианалъной области). У новорожденных детей, в особенности принадлежащих к мошолоидной расе, пигментные клетки дермы часто образуют крупное скопление, располагающееся в области крестца и копчика, которое макроскопически выявляется как пигментированный участок кожи ("монгольское пятно"). Его окраска по интенсивности достигает максимума к 1-му году, а размеры - к 2 годам, после чего оно блекнет и постепенно исчезает (обычно к 6-7 годам). Микроскопически, однако, пигментные клетки в составе дермы определяются у людей любого возраста, более того, в среднем и пожилом возрасте они могут формировать крупные скопления (это нарушение называется меланоцитозом), образуя пигментные пятна - чаще всего в области лица и спины. Содержание пигментных клеток в соединительной ткани
дермы увеличено в области родимых пятен (невусов); оно резко повышено в ней при некоторых заболеваниях кожи, связанных с образованием очагов гиперпигментации.
Численное преобладание пигментных клеток над другими клеточными элементами соединительной ткани характерно для радужки и сосудистой оболочки глаза, где им принадлежит и функционально ведущая роль. Такую ткань называют пигментной и относят к одному из видов соединительных тканей со специальными свойствами (см. главу 11).
МЕЖКЛЕТОЧНОЕ ВЕЩЕСТВО
РЫХЛОЙ ВОЛОКНИСТОЙ СОЕДИНИТЕЛЬНОЙ ТКАНИ
Межклеточное вещество рыхлой волокнистой соединительной ткани состоит из волокон и основного аморфного вещества. Оно является продуктом деятельности клеток этой ткани, в первую очередь, фибробластов.
Функции межклеточного вещества рыхлой волокнистой соединительной ткани:
Ф обеспечение архитектоники, физико-химических и механических свойств ткани;
© участие в создании оптимального микроокружения для деятельности клеток;
© объединение в единую систему всех клеток соединительной ткани и обеспечение передачи информации между ними;
@ воздействие на многочисленные функции различных клеток (пролиферацию, дифференцировку, подвижность, экспрессию рецепторов, синтетическую и секреторную активность, чувствительность к действию различных стимулирующих, ингибирующих и повреждающих факторов и т.п.). Этот эффект может осуществляться путем контактного воздействия компонентов межклеточного вещества на клетки, а также благодаря его способности накапливать и выделять факторы роста.
ВОЛОКНА МЕЖКЛЕТОЧНОГО ВЕЩЕСТВА
РЫХЛОЙ ВОЛОКНИСТОЙ СОЕДИНИТЕЛЬНОЙ ТКАНИ
Волокна, входящие в состав межклеточного вещества рыхлой волокнистой соединительной ткани, относятся к трем основным типам,
- 302 каждый из которых обладает особыми морфологическими, механическими и биохимическими свойствами и выполняет определенную функцию в ткани. Различают: (1) коллагеновые волокна, (2) ретикулярные волокна и (3) эластические волокна.
Коллагеновые волокна
Коллагеновые волокна образованы белками коллагенами, которые получили свое название из-за способности содержащих их тканей при длительном вываривании давать животный клей (от греч. kolla - клей и genes - рождающий).
Коллагены - семейство родственных белков, являющихся наиболее распространенными белками в межклеточном веществе соединительных тканей и во всем организме человека (составляют 25-30% их общего количества). Они придают тканям механическую прочность и выполняют морфогенетическую функцию, влияя на рост, миграцию, дифференцировку, секреторную и синтетическую активность различных клеток. Их молекулы способны собираться в филаменты, фибриллы или образовывать сети, взаимодействующие с другими белками межклеточного вещества.
Молекулы коллагенов состоят из трех скрученных спирально полипептидных нитей - ct-цепей (рис. 10-6), в которых преобладают аминокислоты глицин, пролин, лизин, гидроксипролин и гидроксилизин (последние две образуются в грЭПС из пролин а и лизина, соответственно, в ходе синтеза коллагена). Идентифицировано более 30 вариантов а-це-пей коллагена, различных по химическому строению. Каждая из них кодируется отдельным геном, причем разные ткани характеризуются экспрессией тех или иных комбинаций этих генов. Хотя теоретически комбинации всех вариантов a-цепей могли бы дать более 1000 молекулярных форм коллагена, до настоящего времени обнаружено лишь 19 типов (обозначаемых римскими цифрами - I-XIX), из которых наибольшее значение имеют первые пять (см. ниже). Эти коллагены различаются аминокислотным составом их а-пепей, порядком чередования в них аминокислот, молекулярной массой, распределением в тканях.
Коллагены I, II, III и V типов называются фибриллярными, или интерстициальными, так как они образуют фибриллы, которые входят в состав соединительных тканей; коллаген IV типа относят к аморфным (образует плоские сети).
К
 летки,
вырабатывающие коллагены, помимо
фибробластов,
включают
остеобласты,
хондробласты, одонтобласты, цементоблас-ты,
ретикулярные клетки, гладкие миоциты,
клетки периневрия. Коллагены
IV и V типов (как компоненты базальной
мембраны) продуцируются также
эпителиальными
клетками, адипоцитами, гладкими
миоци-тами, кардиомиоцитами, волокнами
склетной мышечной ткани, клетками
нейроглии. Процессы
биосинтеза коллагена наиболее подробно
изучены применительно к фибробластам,
однако они происходят сходным образом
и в указанных выше клетках.
летки,
вырабатывающие коллагены, помимо
фибробластов,
включают
остеобласты,
хондробласты, одонтобласты, цементоблас-ты,
ретикулярные клетки, гладкие миоциты,
клетки периневрия. Коллагены
IV и V типов (как компоненты базальной
мембраны) продуцируются также
эпителиальными
клетками, адипоцитами, гладкими
миоци-тами, кардиомиоцитами, волокнами
склетной мышечной ткани, клетками
нейроглии. Процессы
биосинтеза коллагена наиболее подробно
изучены применительно к фибробластам,
однако они происходят сходным образом
и в указанных выше клетках.
Биосинтез коллагеновых волокон. Образование коллагеновых волокон включает два этапа: (а) внутриклеточный и (б) внеклеточный (рис. 10-7).
Внутриклеточный этап:
(1) Образование иРНК, кодирующих синтез сс-цепей коллагена, в результате транскрипции соответствующих генов (происходит в ядре фибробласта).
Поглощение и транспорт аминокислот, необходимых для синтеза коллагена, механизмом эндоцитоза.
Синтез полипептидных а-цепей из аминокислот на рибосомах грЭПС (трансляция) под контролем иРНК, поступивших в цитоплазму из ядра. Образуются молекулы с длинными краевыми ("регистрационными") пептидами, которые, как предполагают, необходимы для (а) последующей правильной сборки трех а-цепей в молекулы проколлагена, (б) обеспечения его растворимости в воде и (в) предотвращения самопроизвольной сборки фибрилл внутри клетки. Синтезированные пепи накапливаются в просвете цистерн грЭПС.
Пострансляционные изменения - ферментное гидроксилирова-ние пролина и лизина (зависит от витамина С, играющего роль кофактора ферментов), гликозилировние гидроксилизина, а также образование дисульфидных мостиков - осуществляются в просвете цистерн грЭПС.
Образование молекулы проколлагена в результате сборки (скручивания) трех а-цепей - происходит в просвете грЭПС.
Перенос молекул проколлагена из грЭПС в комплекс Гольджи -осуществляется мембранными транспортными пузырьками.
Терминальное гликозилирование и упаковка молекул проколлагена в секреторные пузырьки - происходят в комплексе Гольджи.
Транспорт молекул проколлагена в секреторных пузырьках, отщепляющихся от комплекса Гольджи, к плазмолемме (обеспечивается элементами цитоскелета - микротрубочками и микрофиламентами).
Экзоцитоз молекул проколлагена в участке инвагинации цитоплазмы фибробласта ("бухточки").
Внеклеточный этап (сборка фибрилл - фибриллогенез):
Отщепление регистрационных пептидов проколлагена с помощью связанных с плазмолеммой специфических протеаз (проколла-ген пептидаз) с образованием нерастворимого тропоколлагена, способного к самосборке в фибриллы.
Полимеризация тропоколлагена с образованием коллагеновых фибрилл и волокон протекает самопроизвольно с участием протеоглика-нов и структурных гликопротеинов, секретируемых фибробластами. Структура фибрилл стабилизируется благодаря формированию ковален-тных мостиков между молекулами тропоколлагена под действием фермента лизил оксидазы, секретируемого фибробластами.
Коллагеновые фибриллы толщиной 20-120 нм, обладающие типичной для коллагенов I, II и III типов поперечной исчерченностью (с периодичностью 64-68 нм), образуются в результате упорядоченной агрегации молекул тропоколлагена (диаметром 1.5 нм). Последние связываются в продольные пепочки, разделяясь небольшими промежутками (см. рис. 10-6). Располагаясь параллельно друг другу, такие цепочки образуют пучки, в которых каждая из молекул тропоколлагена сдвинута по отношению к таковой в соседней цепочке на четверть своей длины. Полагают, что при такой конфигурации молекулы оптимальным образом связывают и усиливают друг друга в пределах коллагеновой фибриллы, не создавая в ней "слабых точек", по которым мог бы происходить ее поперечный разрыв.
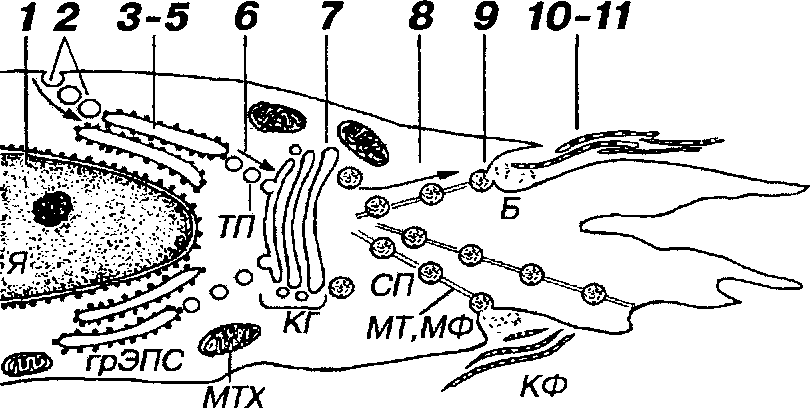
Рис. 10-7. Последовательные этапы синтеза коллагена и образования коллагеновых волокон фибробластом. Внутриклеточный этап: в ядре (Я) происходит образование иРНК, кодирующих синтез а-цепей коллагена (1); аминокислоты, необходимые для синтеза коллагена, поглощаются механизмом эндоцитоза и транспортируются к грЭПС (2); в грЭПС осуществляются процессы синтеза полипептидных а-цепей из аминокислот и их накопление в просвете цистерн (3), пострансляционные изменения а-цепей (4), образование молекулы проколлагена в результате сборки трех а-цепей (5). Молекулы проколлагена переносятся из грЭПС в комплекс Гольджи (КГ) посредством транспортных пузырьков (ТП) - (6); внутри КГ (7) происходит терминальное гликозилирование и упаковка молекул проколлагена в секреторные пузырьки (СП); молекулы проколлагена транспортируются в СП из КГ к плазмолемме (с помощью микротрубочек и микрофиламентов (MT и МФ) - (8) и выделяются экзоцитозом в области "бухточки" (Б) - (9). Внеклеточный этап (фибриллогенез): отщепление регистрационных пептидов проколлагена с образованием нерастворимого тропоколлагена (10); полимеризация тропоколлагена с образованием коллагеновых фибрилл (КФ) и волокон (11). Процессы синтеза и секреции коллагена требуют большого количества энергии, продуцируемой митохондриями (MTX).
Поперечная исчерченность коллагеновых фибрилл (см. рис. 10-6) обусловлена тем, что при так называемом негативном окрашивании краситель заполняет промежутки (зазоры) между молекулами тропоколлагена, которые приобретают вид повторяющихся поперечных темных полос на фибриллах. Линейные участки фибрилл при этом не окрашиваются и имеют вид светлых полос, расположенных между темными. При позитивном окрашивании создается более сложный рисунок поперечной исчерченности коллагеновой фибриллы, включаюпгий несколько полос различной ТОЛПЩНЫ.
Утолщение фибрилл происходит постепенно вследствие присоединения новых молекул тропоколлагена, однако этот процесс находится под контролем, причем для каждого вида ткани характерен свой диаметр фибрилл. В рыхлой волокнистой ткани недавно образованные ("молодые") фибриллы цилиндрической формы во много раз тоньше более зрелых, часто имеющих неправильную форму.
Коллагеновые протофибриллы и коллагеновые микрофибриллы -промежуточные уровни организации коллагеновых структур (между уровнем молекул тропоколлагена и коллагеновых фибрилл), выделяемые рядом авторов. Коллагеновые протофибриллы диаметром 3-5 нм образованы пучками молекул тропоколлагена; коллагеновые микрофибриллы диаметром до 20 нм образованы несколькими протофибриллами.
Объединение коллагеновых фибрилл в пучки приводит к формированию коллагеновых волокон толщиной 1-20 мкм (см. рис. 10-6).
Разрушение (деградация) коллагена фибробластами осуществляется двумя основными путями - внутриклеточным и внеклеточным.
Внутриклеточное разрушение коллагена может происходить до и после секреции синтезированного материала. От 10 до 50% вновь образованного коллагена разрушается самим фибробластом в течение ближайшего времени после его продукции. При этом значительная часть коллагена после синтеза не выделяется из клетки. Предполагают, что такая внутриклеточная деградация коллагена связана с активностью некоего механизма "контроля качества", который выявляет и уничтожает молекулы с измененной структурой, возникшие вследствие "биологических ошибок". Фибробласты подвергают внутриклеточному разрушению и коллагеновые фибриллы, находящиеся в межклеточном пространстве. Для этого они сначала распознают фибриллу, которая подлежит разрушению, затем частично расщепляют ее путем ферментного воздействия, фагоцитируют ее фрагменты и переваривают их внутриклеточно с помощью лизосомальных ферментов.
Внеклеточное разрушение коллагена осуществляется путем секреции фибробластами в межклеточное пространство группы ферментов (из которых наиболее изучена коллагеназа), обеспечивающих внеклеточное расщепление белков межклеточного вещества до мелких пептидных фрагментов.
Коллагеновые волокна толщиной 1-20 мкм образуются путем объединения в пучки коллагеновых фибрилл (см. рис. 10-6). В рыхлой волокнистой соединительной ткани коллагеновые волокна (собственно коллагеновые волокна) образованы преимущественно коллагеном I типа. На препаратах они имеют вид оксифилыгых продольно исчерченных извитых тяжей, идущих в различных направлениях поодиночке и часто образующих пучки вариабельной (до 150 мкм) толщины - (см. рис. 10-1). При изучении в поляризационном микроскопе обнаруживается, что коллагеновые волокна обладают свойством двойного лучепреломления, что указывает на наличие продольно расположенных субмикроскопических единиц. При исследовании под электронным микроскопом выявляются образующие их параллельно лежащие фибриллы диаметром 20-120 нм с поперечной исчерченностью (период 64-68 нм) -см. рис. 10-6.
Основные функции коллагеновых волокон:
Ф обеспечение высоких механических свойств соединительной ткани. Чем выше содержание коллагеновых волокон в данной ткани, тем большей прочностью она обладает. Эти волокна практически нерастяжимы; при увеличении нагрузки они лишь слегка распрямляются, утрачивая волнообразный ход и более не удлиняясь вплоть до достижения предела прочности, превышение которого вызывает их разрыв;
© определение (в значительной мере) архитектоники соединительной ткани;
© обеспечение взаимодействий между клетками и межклеточным веществом, а также связь между отдельными компонентами межклеточного вещества;
@ влияние на пролиферацию, дифференцировку, миграцию и функциональную активность различных клеток.
Нарушения синтеза коллагена и сборки коллагеновых волокон очень многообразны и могут явиться результатом дефектов (обычно генетически обусловленных) отдельных стадий внутриклеточного или внеклеточного этапов их образования. Они лежат в основе ряда заболеваний, связанных с мутациями генов, кодирующих молекулы коллагена. К таким заболеваниям, в частности, относятся различные формы синдрома Элерса-Данло (Ehlers-Danlos), при котором у больных отмечаются повышенная эластичность кожи, патологическая подвижность суставов, разрывы стенки аорты и (или) кишки; различные формы несовершенного остеогенеза (osteogenesis imperfecta), для которого характерны патологическая ломкость костей и нарушения функции сердца (см. также главу 12).
Поскольку для синтеза коллагена необходим витамин С (аскорбиновая кислота), его недостаточное поступление в организм вызывает серьезные расстройства (цингу), проявляющиеся, наряду с другими симптомами (мышечной слабостью, кровоточивостью, отечностью и изъязвлением десен), признаками, обусловленными нарушением выработки коллагена. В частности, при цинге происходит расшатывание и выпадение зубов (из-за нарушения обновления волокон периодонтальной связки - главного элемента поддерживающего аппарата зуба), замедляется заживление ран и костных переломов.
Нарушение баланса между образованием и разрушением коллагена может приводить к избыточному отложению коллагеновых волокон (диффузному или очаговому), которое характерно для фиброза - патологического состояния, возникающего в различных органах (печени, почках, легких, миокарде) и вызывающего нарушение их функций. Избыточное накопление коллагена в участках повреждения кожи приводит к формированию утолщенных келоидных рубцов (от греч. kele -опухоль и oidos - подобный).
Ретикулярные волокна
Ретикулярные волокна имеют малый диаметр (0.1-2 мкм) и, как правило, формируют тонкие растяжимые трехмерные сети, что определило их название (от лат. reticulum - сеточка). Они образованы коллагеном III типа, т.е. по своей химической природе также являются колла-геновыми. Эти волокна не обнаруживаются на препаратах, окрашенных гематоксилином и эозином. Их выявление основано на способности давать ШИК-реакцию и окрашиваться солями серебра, отчего их называют также аргирофильными (от греч. argyros - серебро и philia - любовь). Каждое ретикулярное волокно образовано пучком микрофибрилл толщиной 20-40 нм, обладающих поперечной исчерченностью с периодичностью 64-68 нм и заключенных в оболочку из гликопротеинов и протеогликанов, которая и обусловливают аргирофилию и ШИК-реакцию этих волокон.
Основная функция ретикулярных волокон - опорная Они встречаются в рыхлой волокнистой соединительной ткани (особенно во вновь образованной или подвергающейся перестройке), а также во всех других видах соединительной ткани. Ретикулярные волокна многочисленны в кроветворных (миелоидной и лимфоидной) тканях, в которых вместе с ретикулярными клетками образуют поддерживаюпгдй каркас для развивающихся элементов крови (см. рис. 9-8). Ретикулярные во-
-310локна входят в состав базальных мембран (образуя их ретикулярную пластинку), располагаются между эпителиальными структурами в печени и почке, окружают капилляры и нервные волокна.
Клетки, обладающие способностью к выработке ретикулярных волокон, помимо фибробластов включают ретикулярные и жировые клетки, гладкие миоциты, кардиомиоциты, нейролеммоциты (клетки, образующие оболочку нервных волокон в периферической нервной системе). Эта способность характерна и для симпластических образований - волокон скелетной мышечной ткани.
Эластические волокна
Эластические волокна в соединительной ткани обычно содержатся в значительно меньшем количестве, чем коллагеновые, за исключением участков, обладающих подвижностью. На светооптическом уровне они выявляются при использовании избирательных методов окраски (чаще всего - орсеина). Эластические волокна варьируют по толщине в пределах 0.2-10 мкм, ветвятся и анастомозируют друг с другом, формируя трехмерные сети (см. рис. 10-1); в отличие от коллагеновых волокон, они обычно не образуют пучки.
Функции эластических волокон: © определение архитектоники ткани;
© обеспечение способности ткани к обратимой деформации (к возвращению к исходной форме после ее временного изменения.
Эластин - главный белковый компонент эластических волокон. Он составляет более 90% их массы и представлен гликопротеиновыми молекулами, имеющими в состоянии покоя форму скрученных нитей. При растяжении они распрямляются, а после прекращения действия нагрузки - вновь закручиваются. Молекулы эластина ковалентно "сшиты" друг с другом в комплексы, формирующие эластические волокна и пластинки (мембраны).
Структурные компоненты эластических волокон выявляются на электронно-микроскопическом уровне. Каждое волокно содержит: (а) центральный светлый (аморфный) компонент, образованный эластином, (б) периферический (микрофибриллярный) компонент, состоящий из волоконец толщиной 10-12 нм, образованных гликопротеином фибриллином. Элементы микрофибриллярного компонента частично погружены в аморфный компонент (рис. 10-8).
-
311 
отв элв зэв
Эластическая система - совокупность волокон, обладающих эластическими свойствами. Помимо собственно эластических волокон, являющихся ее основным и наиболее зрелым элементом, к ней относят также окситалановые и элауниновые волокна. Первые образованы микрофибриллами толщиной 10-12 нм, сходными с теми, которые окружают центральный аморфный компонент эластических волокон, вторые по строению занимают промежуточное положение между типичными эластическими и окситалановыми (см. рис. 10-8).
Синтез и взаимосвязь элементов эластической системы. Микро-фибриллярный компонент, первоначально синтезируемый фибробластами, как предполагают, служит структурной основой, на которую далее эти клетки откладывают эластин. Поэтому, по мере созревания эластического волокна, эластин постепенно накапливается в его центральной части, а микрофибриллярный компонент оттесняется к периферии волокна и в конечном итоге почти полностью разрушается. Таким образом, формирование эластического волокна (эластогенез) описывается последовательностью:
окситалановое волокно —> элауниновое волокно -> эластическое волокно (см. рис. 10-8).
В соответствии с этой схемой окситалановые и элауниновые волокна можно рассматривать как незрелые эластические.
Клетки, вырабатывающие эластические волокна (помимо фиб-робластов) включают: гладкие миоциты, хондробласты и хондроциты. Микрофибриллы входят в состав межклеточного вещества мезангия в почечном клубочке, образуют волокна ресничного пояска (цинновой связки), удерживающие хрусталик.
Структурные изменения эластических волокон, обусловливающие нарушение их функциональных свойств, выявлены при ряде заболеваний, связанных с мутациями генов, кодирующих синтез соответствующих белков. У таких больных выявляется ненормальная растяжимость кожи, повышенная подвижность суставов, аномалии сердца и сосудов. При синдроме Марфана выявлено нарушение синтеза фибрил-лина (микрофибриллярного компонента эластических волокон). Такие больные погибают в возрасте до 35 лет (при неонатальной форме -в раннем детстве) преимущественно вследствие аномалий органов сердечно-сосудистой системы, неспособных выдерживать нормальные функциональные нагрузки (наиболее часто - вследствие разрыва аорта). Для этого синдрома характерны также изменения кожи, суставов, скелета, смещение хрусталика.
ОСНОВНОЕ АМОРФНОЕ ВЕЩЕСТВО
РЫХЛОЙ ВОЛОКНИСТОЙ СОЕДИНИТЕЛЬНОЙ ТКАНИ
Основное аморфное вещество заполняет промежутки между волокнистыми компонентами межклеточного вещества и окружает клетки. При изучении под светооптическим и электронным микроскопами оно имеет аморфное строение, прозрачно, характеризуется базофили-ей и низкой электронной плотностью. На молекулярном уровне оно обладает сложной организацией и состоит из макромолекулярных гид-ратированных комплексов протеогликанов (см. рис. 12-3) и структурных гликопротеинов.
Протеогликаны состоят из пептидной цепи, связанной с гликоз-сшиногликанами (ГАГ). Строение молекулы протеогликанов описано в главе 12.
Гликозаминогликаны (ГАГ) - крупные неразветвленные отрицательно заряженные гидрофильные полисахаридные молекулы, образованные повторяющимися дисахаридными единицами. Основными ГАГ в организме человека являются: гиолуроновая кислота, хондроитин-сульфат, дерматансульфат, гепарансульфат и гепарин, а также кера-тансульфат. За исключением гиалуроновой кислоты, ГАГ связываются с белками, образуя протеогликаны. Присутствие определенных типов ГАГ в различных тканях определяет свойства их межклеточного вещества, в частности, его проницаемость и способность связывать другие молекулы.
Протеогликаны синтезируются в грЭПС и комплексе Гольджи фиб-робластов, после чего выделяются механизмом экзопитоза в межклеточное пространство, где они, вероятно, объединяются в крупные протео-гликановые агрегаты. Обновление протеогликанов в тканях происходит более интенсивно, чем коллагена. Они разрушаются рядом лизосомаль-ных ферментов клеток соединительной ткани; при дефектах или недостаточности этих ферментов развиваются заболевания, обусловленные накоплением в клетках частично переваренных протеогликанов - муко-полисахаридозы.