

Гистогенетическая классификация эпителиев (по н.Г.Хлопину)
Гистогенетический тип эпителия |
Эмбриональные зачатки -источники развития эпителия |
1. Эпидермальный 2. Энтеродермальный 3. Целонефродермальный 4. Ангиодермалъный 5. Эпендимоглиальный |
эктодерма, прехордальная пластинка кишечная энтодерма целомическая выстилка, нефротом ангиобласт нервная трубка |
/. Эпителии эпидермального типа развиваются из эктодермы и прехордальной пластинки, которая также обладает эктодермальной детерминацией. Самым типичным представителем данного тканевого типа служит эпителий кожи (эпидермис), наиболее общей функцией - защитная, самым характерным морфологическим признаком - многослой-ность или многорядность (последняя при повреждении эпителия может сменяться многослойностью).
К эпидермальному типу помимо эпидермиса относят эпителии, выстилающие полость рта, глотки, пищевода, воздухоносных путей и респираторного отдела легких, мочевыводящих путей, влагалища, роговицу глаза. Эпителии эпидермального типа образуют железы кожи и железы, связанные с указанными выше слизистыми оболочками, функционально ведущую ткань крупных слюнных и молочных желез, ряда эндокринных желез (аденогипофиза, щитовидной и околощитовидных желез), эпителиальную основу тимуса. Некоторые авторы относят эпителии - производные прехордальной пластинки не к эпидермальному, а к энтеродер-мальному тканевому типу.
Эпителии энтеродермалъного типа (от греч. еШегоп - кишка) являются производными кишечной энтодермы. Самым типичным представителем этих эпителиев служит эпителий кишки, наиболее типичными функциями - всасывание и (или) секреция, самым характерным морфологическим признаком - однослойность. Эпителии энтеродермалъного типа образуют выстилку желудочно-кишечного тракта, а также все связанные с ней железы (от самых мелких, расположенных в его стенке, до самых крупных - печени и поджелудочной железы).
Эпителии целонефродермального типа (от греч. сое1от - целом, полость тела и перЬгоБ - почка) развиваются из целомической выстилки и нефротома. Их строение весьма разнообразно; наиболее часто они образуют однослойные однорядные выстилки, но встречаются также однослойные многорядные (например, в семявыносящих путях, предстательной железе) и многослойные (например, в фолликулах яичника). Функции этих эпителиев связаны с процессами секреции, экскреции и всасывания; они выполняют также барьерную функцию. К эпителиям целонефродермального типа относят мезотелий (однослойный плоский эпителий, образующий выстилку полостей тела и входящий в состав серозных оболочек), эпителий нефрона, семявыносящих путей, выстилки маточных труб, матки, шеечного канала. Эти эпителии входят в состав яичника (фолликулярные клетки, зернистые лютеоциты) и яичка (сустентоциты, или клетки Сертоли), образуют функционально ведущую ткань предстательной железы и коркового вещества надпочечника.
Эпителий ангиодермального типа (от греч. ап^оп - сосуд) являются производными особого эмбрионального зачатка ангиобласта, который располагается среди клеток мезенхимы и дает начало выстилке кровеносных и лимфатических сосудов, а также сердца. Единственным представителем данного типа служит эндотелий, который обычно является однослойным плоским эпителием. Исключение составляют выстилки сосудов с особыми функциональными свойствами - (а) посткапиллярных венул с высоким (кубическим) эндотелием в органах иммунной системы, (б) синусов селезенки с палочковидными эндотелиальными клетками. Функции эндотелия многообразны: он участвует в процессах двустороннего транспорта веществ и клеток между кровью и другими тканями, регуляции свертывания крови и тонуса сосудов, секретирует биологически активные вещества, обеспечивает новообразование сосудов (ангиогенез).
5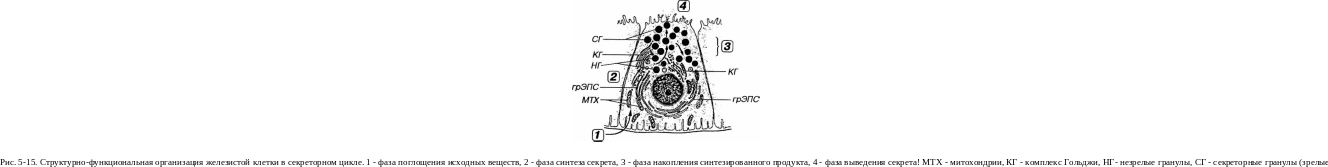 .
Эпителии
эпендимоглиального типа - особые
ткани нейраль-ного происхождения,
выполняющие в нервной системе опорную,
разграничительную и секреторную
функции, имеющие строение эпителия и
относящиеся к так называемой эпендимной
глии, давшей
им свое название (от греч. ependyma
- верхняя одежда и glia,
или neuroglia
-нервный клей, т.е. ткань, связывающая
элементы нервной системы). Этот эпителий
выстилает центральный канал спинного
мозга и желудочки головного мозга,
участвуют в выработке спинномозговой
жидкости (см. главу 14). Он образован
одним слоем кубических или призматических
клеток, которые часто имеют реснички
на апикальной поверхности и длинный
базалыгым отросток. По мнению некоторых
авторов, эпителии эпендимоглиального
типа образуют также выстилки мозговых
оболочек, входят в состав органов чувств
(равновесия, слуха, зрения) и, возможно,
нервных стволов (формируя периневрий).
.
Эпителии
эпендимоглиального типа - особые
ткани нейраль-ного происхождения,
выполняющие в нервной системе опорную,
разграничительную и секреторную
функции, имеющие строение эпителия и
относящиеся к так называемой эпендимной
глии, давшей
им свое название (от греч. ependyma
- верхняя одежда и glia,
или neuroglia
-нервный клей, т.е. ткань, связывающая
элементы нервной системы). Этот эпителий
выстилает центральный канал спинного
мозга и желудочки головного мозга,
участвуют в выработке спинномозговой
жидкости (см. главу 14). Он образован
одним слоем кубических или призматических
клеток, которые часто имеют реснички
на апикальной поверхности и длинный
базалыгым отросток. По мнению некоторых
авторов, эпителии эпендимоглиального
типа образуют также выстилки мозговых
оболочек, входят в состав органов чувств
(равновесия, слуха, зрения) и, возможно,
нервных стволов (формируя периневрий).
ЖЕЛЕЗЫ
Железы выполняют секреторную функцию, вырабатывая и выделяя разнообразные продукты (секреты), обеспечивающие различные функции организма. Большинство желез образовано эпителиальной тканью (железистым эпителием), хотя той или иной способностью к секреции обладают все ткани.
митохондрий, находящихся в цитоплазме. Избыток синтезируемых продуктов часто удаляется внутриклеточным механизмом кринофагии, что обусловлено хорошим развитием лизосомального аппарата. Распределение органелл в цитоплазме клеток желез неравномерно в связи с их выраженной полярностью.
Строение и гистофизиология желез
Железистые клетки (гландулоциты - от лат. glándula - железа и kytos, или cytos - клетка) специализированы на выработке секретов, поэтому для них характерны все признаки клеток с активно протекающими синтетическими процессами (рис. 5-15).
Ядро гландулоцитов - обычно крупное, с преобладанием эухрома-тина, одним или несколькими крупными ядрышками. Его положение в клетке может изменяться в разные фазы секреторного цикла (см. ниже), например, оно может смещаться к базальному полюсу при накоплении секреторных гранул в апикальном.
Цитоплазма гландулоцитов содержит мощно развитый синтетический аппарат, морфологические и функциональные особенности которого зависят от химической природы продуцируемого секрета (см. главу 3). Процессы синтеза и выделения веществ требуют значительного количества энергии, которая вырабатывается большим числом
Секреторный цикл. Процесс секреции в железистых клетках протекает циклически и включает четыре фазы, которые могут в различной степени взаимно перекрываться: (I) фазу поглощения исходных веществ, (2) фазу синтеза секрета, (3) фазу накопления синтезированного продукта, (4) фазу выведения секрета (см. рис. 5-15).
1. Фаза поглощения исходных веществ, служащих субстратами для синтеза секреторного продукта, обеспечивается высокой активностью транспортных механизмов, связанных с плазмолеммой базаль-ного полюса клетки, через который указанные вещества поступают из крови. В некоторых клетках субстраты для синтеза могут в значительных количествах запасаться в цитоплазме (например, в виде липидных капель в стероид-продуцирующих клетках).
Фаза синтеза секрета связана с процессами транскрипции и трансляции, деятельностью грЭПС и комплекса Гольджи (для белковых секретов), аЭПС и митохондрий с тубулярно-везикулярными крис-тами (для стероидных веществ). Синтезированный продукт в комплексе Гольджи или внутри секреторных гранул нередко претерпевает посттрансляционные изменения, обусловленные действием различных ферментов ("дозревает").
Фаза накопления синтезированного продукта в цитоплазме железистых клеток обычно проявляется нарастанием содержания секреторных гранул, которые в некоторых случаях могут укрупняться, сливаясь друг с другом. Переполнению цитоплазмы секреторными гранулами препятствует механизм лизосомального разрушения их избытка - крино-фагия. Скопления гранул располагаются преимущественно у апикального полюса клеток экзокринных желез и у базального - в клетках эндокринных желез (см. ниже). Некоторые виды синтезированных продуктов (например, стероидные гормоны) не накапливаются в цитоплазме железистых клеток, а по мере образования, по-видимому, сразу же из нее выводятся.
Фаза выведения секрета может осуществляться несколькими механизмами (см. ниже). Наиболее часто происходит экзоцитоз содержимого секреторных гранул путем слияния мембраны их гранул с плаз-молеммой и выделения синтезированного продукта за пределы клетки. Встроенная в плазмолемму мембрана секреторных гранул затем отделяется из нее в цитоплазму механизмом эндоцитоза и возвращается в комплекс Гольджи для повторного использования (реутилизации, или рециклирования). Некоторые секреты (например, стероидные или тире-оидные гормоны) выделяются из клетки механизмами диффузии.
Классификация желез
Существует несколько классификаций желез, которые основаны на учете различных признаков. Железы подразделяются:
по числу клеток (рис. 5-16) - на одноклеточные (например, бокаловидные клетки, клетки диффузной эндокринной системы) и многоклеточные (большинство желез);
по уровню организации - на входящие в состав различных органов в качестве их компонентов (например, железы слизистых оболочек) или являющиеся самостоятельными анатомически оформленными органами (например, крупные слюнные железы, печень, поджелудочная железа, щитовидная железа и др.);
3) по расположению (относительно эпителиального пласта) - на эндоэпителиальные и экзоэпителиальные, т.е. лежащие в пределах эпителиального пласта или вне его, соответственно (см. рис. 5-16). Большинство желез относится к экзоэпителиальным;
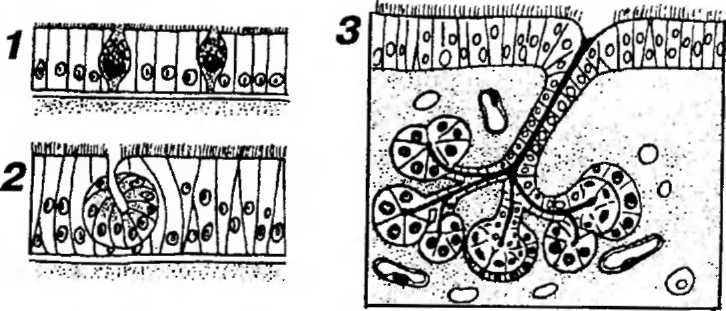
Рис. 5-16. Классификация экзокринных желез в зависимости от количества клеток, входящих в их состав, и расположения по отношению к эпителиальному пласту. 1 - одноклеточные эндоэпителиальные железы (бокаловидные клетки);
- многоклеточная эндоэпителиальная железа (слизистой оболочки полости носа);
- многоклеточная экзоэпителиальная железа (слизистой оболочки трахеи или бронхов).
по месту (направлению) выведения секрета - на эндокринные (вьщеляющие секреторные продукты, называемые гормонами, в кровь) и экзокринные (вьщеляющие секреты на поверхность тела или в просвет внутренних органов);
по механизму (способу) выведения секрета (рис. 5-17) - на мерокринные (без нарушения структуры клетки), апокринные (с отделением в секрет части апикальной цитоплазмы) и голокринные (с полным разрушением клеток и выделением их фрагментов в секрет). В организме человека большинство желез относится к мерокринным; апокринных желез немного (например, часть потовых и молочные), к голокринным относятся лишь сальные железы. В клетках некоторых желез выведение секрета осуществляется одновременно двумя механизмами - апокрин-ным и мерокринным.
6) по химическому составу вырабатываемого секрета - на
белковые (серозные), слизистые, смешанные (белково-слизистые), ли-пидные и др. Характеристики (5) и (6) используются преимущественно в отношении экзокринных желез.
Э ндокринные
железы (железы внутренней секреции)
продуцируют
гормоны
- вещества
различной
химической
природы, циркулирующие в крови в
низких концентрациях и обладающие
высокой биологической активностью.
Эндокринные железы имеют различное
строение и уровень организации - от
одноклеточных
(элементы
диффузной эндокринной системы) до
сравнительно крупных
органных образований (например,
щитовидная железа, надпочечники,
гипофиз, эпифиз). Синтезированные
клетками эндокринных желез продукты
(гормоны) выводятся через базальный
полюс клетки; выводные
протоки в таких железах отсутствуют.
ндокринные
железы (железы внутренней секреции)
продуцируют
гормоны
- вещества
различной
химической
природы, циркулирующие в крови в
низких концентрациях и обладающие
высокой биологической активностью.
Эндокринные железы имеют различное
строение и уровень организации - от
одноклеточных
(элементы
диффузной эндокринной системы) до
сравнительно крупных
органных образований (например,
щитовидная железа, надпочечники,
гипофиз, эпифиз). Синтезированные
клетками эндокринных желез продукты
(гормоны) выводятся через базальный
полюс клетки; выводные
протоки в таких железах отсутствуют.
Экзокринные железы вырабатывают разнообразные по химической природе и функциональному значению секреты и так же, как и эндокринные железы, различаются по строению и уровню организации. В экзокринных железах выделяют (1) концевые (секреторные) отделы и (2) выводные протоки.
Концевые (секреторные) отделы состоят из железистых клеток, которые продуцируют секрет. В некоторых железах, образованных эпителия ми эпидермального типа (например, потовых, молочных, слюнных), концевые отделы помимо железистых клеток содержат особые отростчатые миоэпителиальные клетки - видоизмененные эпители-оциты с развитым сократительным аппаратом. Миоэпителиальные клетки своими отростками охватывают снаружи железистые и, сокращаясь, способствуют выведению секрета из концевого отдела.
Выводные протоки связывают концевые отделы с покровными эпителиями и обеспечивают выделение синтезированных продуктов на поверхность тела или в полость органов. Как правило, их клетки не обладают секреторной функцией, хотя могут влиять на конечный состав вьшодимого секрета, в частности, изменяя содержание ионов и воды (например, в потовых и слюнных железах). Мелкие протоки отдельных желез могут содержать миоэпителиальные клетки (в тех случаях, когда они имеются в концевых отделах). Во многих крупных железах выводные протоки образуют сложную систему, различные участки которой выполняют специализированные функции и обладают неодинаковым строением.
Разделение на концевые отделы и выводные протоки затруднено в некоторых железах (например, желудка, матки), так как все их клетки обладают свойствами секреторных.
Морфологическая классификация экзокринных желез основана на структурных признаках их концевых отделов и вьшодньгх протоков. Железы подразделяются (рис. 5-18):
по форме концевых отделов - на трубчатые и альвеолярные (сферические); при наличии обеих форм железы называются трубчато-альвеолярными или альвеолярно-трубчатыми;
по ветвлению концевых отделов - на неразветвленные и разветвленные;
по ветвлению выводных протоков - на простые (с неразвет-вленным протоком) и сложные (с разветвленными протоками).
Р
 азвитие
эндокринных и экзокринных желез на
начальных этапах осуществляется
сходным образом - путем формирования
покровным эпителием тяжа, внедряющегося
в
подлежащую мезенхиму
(рис.
5-19), которая оказывает на него индуцирующее
воздействие.
В дальнейшем этот тяж растет (и часто
ветвится) вследствие интенсивного
деления его клеток.
азвитие
эндокринных и экзокринных желез на
начальных этапах осуществляется
сходным образом - путем формирования
покровным эпителием тяжа, внедряющегося
в
подлежащую мезенхиму
(рис.
5-19), которая оказывает на него индуцирующее
воздействие.
В дальнейшем этот тяж растет (и часто
ветвится) вследствие интенсивного
деления его клеток.
В экзокринных железах эпителиальные клетки, расположенные в дистальных участках этого тяжа, дифференцируясь, приобретают признаки секреторных клеток и формируют концевые (секреторные) отделы. Эпителиальные клетки проксимальной части тяжа образуют выводные протоки - систему трубочек, связывающих концевые отделы с покровным эпителием в области начального формирования закладки железы.
В эндокринных железах клетки дистальной части эпителиального тяжа дифференцируются в секреторные и вступают в связь с многочисленными формирующимися сосудами; проксимальная часть тяжа разрушается, вследствие чего эндокринная железа утрачивает связь с покровным эпителием, давшим начало ее закладке. При аномалиях развития такая связь может сохраняться, а проксимальная часть эпителиального тяжа - формировать подобие выводного протока (например, известны аномалии развития щитовидной железы, при которых она сохраняет связь с местом своей закладки - слепым отверстием языка - посредством ductus thyreoglossus).