

В) Особенности структурной организации передних отделов коры
Первичные, вторичные и третичные поля затылочной, височной и теменной коры, о которых до сих пор шла речь, сконцентрированы в задней части полушарий, располагающейся позади центральной борозды. Расположенные кпереди от этой борозды поля лобной доли мозга, включающей в себя прецентральную и собственно лобную области, характеризуются некоторыми функциональными и структурными чертами, существенно отличными от особенностей той части коры, которая находится позади центральной борозды.
В составе лобной доли мозга мы можем выделить принципиально те же три основные группы полей, что и в рассмотренных выше областях.
Прецентральная область, непосредственно граничащая спереди с постцентральной областью, еще со времен Фрича и Гитцига известна нейрофизиологам и клиницистам как двигательная зона коры. В ходе филогенеза эта область дифференцируется из единой сензомоторной коры и распадается на два поля: первичное гигантопирамидное, или моторное, поле 4 (центральное двигательное поле) и вторичные премо-торные поля 6 и 8 (периферические двигательные поля). Общая структурно-функциональная характеристика этих полей и их связей с подкорковыми отделами мозга обнаруживает выраженные черты сходства с первичными и вторичными полями затылочной, височной и постцентральной коры. Вместе с тем имеются, как уже сказано, и существенные различия, которые обусловлены тем, что в организации коры всей лобной доли мозга, в отличие от задней части полушария, основной акцент падает на осуществление координированных и целенаправленных воздействий организма на внешний мир в ответ на воспринимаемые комплексы раздражений.
Наиболее бросающимися в глаза особенностями микроскопического строения первичного гигантопирамидного поля 4 взрослого человека является, как известно, отсутствие в нем отчетливо обособленного слоя IV и наличие мощно развитых гигантских клеток Беца в слое V. Последние дают начало наиболее крупным волокнам пирамидного пути, проводящим импульсы произвольных движений на двигательные центры скелетной мускулатуры, расположенные в стволе головного мозга и в спинном мозгу. Обе эти структурные особенности с несомненностью свидетельствуют о том, что в центральном двигательном поле сравнительно наибольшего развития достигают элементы нейронного комплекса, приспособленные для максимально быстрого и кратчайшим путем проведения произвольных импульсов на эффекторные нейроны центральной нервной системы. Напротив, те элементы корковой организации, которые приспособлены для восприятия афферентных импульсов из подкорки и которые в полях задних отделов полушария в большом количестве сконцентрированы в слое IV, имеют в гиганто пирамидном поле более рассеянное расположение, не образуя четко отграниченного слоя.
Из этого можно заключить, что сильно развитые в поле 4 проекционные афферентные волокна, приносящие импульсы из подкорковых образований, переключают большую часть поступающих импульсов на гигантские эфферентные пирамиды слоя V. Такая конструкция особенно благоприятствует быстрой «отдаче» на двигательную периферию тела всех сигналов, приходящих в данное место коры из подкорковых уровней центральной нервной системы, так же как и из других отделов коры.
Указанные особенности микроскопической структуры, характеризующие гигантопирамидное поле 4 как центральное, как нельзя лучше согласуются с его физиологической и клинической характеристикой, сводящейся, как известно, к весьма дробно дифференцированному у человека представительству в этом поле движений отдельных мышечных групп.
Вторичное премоторное поле 6, как можно судить на основании физиологических и клинических данных (см. II, 4, д), имеет наиболее существенное значение для выполнения и автоматизации более сложных координированных двигательных комплексов, протекающих во времени и вовлекающих в совместную деятельность различные группы мышц тела. При наличии сходства с гигантопира-мидным полем поле б цитоархи-тектонически характеризуется отсутствием гигантских клеток Беца и лучше развитыми крупными пирамидами слоя III (специально его глубокого подслоя). Проекционные связи премоторной коры с подкорковыми образованиями составляют важную часть экстра-пирамидных систем коры, которые в отличие от прямого пирамидного пути достигают конечных двигательных центров головного и опийного мозга через ряд переключений в лежащих ниже коры уровнях центральной нервной системы (рис. 13).
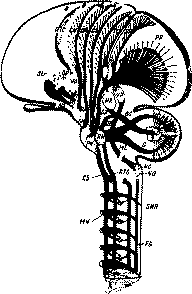
Рис. 13. Схема пирамидной и экстрапирамидных систем (по Пейпецу) :FG—NG—ML—VP—SR — проводящие пути и переключения кожно-кинестетических рецепций: FF/PP} D—BC—NR—VL—TC —связи коры мозга с мозжечком; CS—MN— корково-двигательный пирамидный путь; CS—RN—RTS — связи коры с ядрами ретикулярной формации; CR—SN — связи коры с черным веществом; Str — подкорковые узлы больших полушарий
Глазодвигательное поле 8, также дающее начало одной из экстрапирамидных систем проекционных связей коры, может быть отнесено к периферическому (вторичному) отделу двигательной зоны коры, хотя по своему цитоархитектоническому строению эта формация и обнаруживает уже известные признаки переходности к полям собственно лобной 'коры. Поле 8 имеет отношение к осуществлению координированных движений взора при фиксации внимания и манипулировании с предметами под контролем зрения. На особенностях нарушения движений взора при подобных поражениях мы еще остановимся ниже.
Моторное и премоторные поля прецентральной области, входящие в состав единого кинетического комплекса коры, имеют хорошо развитую проекционную и ассоциационную афферентацию, представленную в виде двусторонних связей, объединяющих эти поля между собой, с другими полями коры и с подкорковыми образованиями. Однако потоки афферентных импульсов, поступающих в поля двигательной коры, имеют источники, отличные от источников поступления импульсов в ядерные зоны анализаторов и в межъядерные зоны коры задней части полушария (рис. 14). В последней собираются, как уже сказано, основные проводники рецепций, идущих от органов чувств на кору, образующие магистральные пути систем анализаторов с их дополнительными переключениями в ближайшей к коре подкорке (определенные ядра зрительных бугров и коленчатые тела).
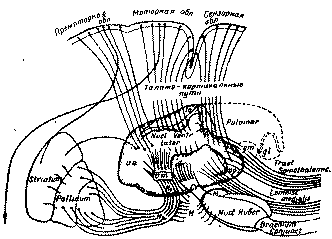
Рис 14 Схема различных систем афферентаций чувствительных и двигательных (кинестетических и кинетических) отделов коры (по Пейпецу и др )
Афферентация, направляющаяся в моторную и премоторную кору, переключается в иной группе ядер зрительного бугра, чем та афферентация, которая поступает в корковые формации задней части полушария. Каналами передачи импульсов, достигающих полей прецентральной области, являются главным образом пути, ведущие от мозжечка через красное ядро и зрительный бугор на кору (рис. 14). Эта афферентация в значительной своей части служит для обратного замыкания тех потоков импульсов, которые циркулируют по экстрапирамидным корково-подкорковым системам мозга (кора — подкорковые узлы больших полушарий— зрительный бугор — кора; кора — мозжечковая система — зрительный бугор — кора; пути поверхностной и глубокой чувствительности — зрительный бугор — подкорковые узлы больших полушарий — зрительный бугор — кора).
Своеобразие качественного состава проекционной афферентаций, обслуживающей двигательную кору, имеет весьма существенное значение для правильной интерпретации функциональной роли этой коры в совокупной деятельности корковых концов анализаторов. Выше мы упоминали о том, что механизм обратной связи находит универсальное применение в рефлекторной деятельности всей центральной нервной системы и специально коры мозга. Однако в ядерных зонах анализаторов с их зонами перекрытия принцип обратной связи реализуется главным образом в процессе восприятия анализаторами различных по сложности раздражений, поступающих из окружающего мира. Физиологическим механизмом этой функции является установка и настройка соответствующих анализаторов на адекватное отражение изменчивой картины действительности.
В деятельности двигательной коры, которая является ответственной за организацию, «программирование» и осуществление произвольной двигательной активности организма, принцип обратной связи получил иное конкретное применение, выступая здесь в качестве основного источника информации об эффекте совершаемых движений и действий. Физиологическая роль двигательной коры в наиболее существенных своих чертах сводится, по-видимому, к сопоставлению «заданной программы» двигательного акта, формирующейся главным образом на основе анали-тико-синтетической работы коры задних отделов полушарий, с реальным ходом выполнения, т. е. к определению сигналов успеха и сигналов ошибок (согласования и рассогласования между программой и действием) и к внесению необходимых и своевременных коррекций по ходу реализуемых действий (А. А. Бернштейн, 1947; Прибрам, 1959, 1960 и др.). Ввиду вышесказанного становится понятным, что как центробежные, так и центростремительные (осуществляющие обратную связь) цепи переключен ий импульсов, соединяющие двигательную зону с подкорковыми образованиями, включены в экстрапирамидные системы мозга, которые, как известно, имеют весьма существенное значение в координированном протекании всех произвольных движений.
Разделение двигательной зоны коры на моторное поле и премотор-ные поля возникло в результате прогрессивной структурной дифферен-цировки в эволюции первоначально единой корковой закладки по мере усложнения корково обусловленных активных двигательйых возможностей организма. Дальнейшее развитие и совершенствование всей сферы так называемых произвольных, направленных во внешний мир актов, было связано с формированием третичных полей собственно лобной области. Эти поля, занимая у человека около 'А всей протяженности коры, относятся к филогенетически наиболее молодым разделам новой коры. Они сформировались в результате разрастания кпереди, по направлению к лобному полюсу, элементов того типа нейронной организации, который заложен в основе двигательной зоны коры.
В лобной области обнаруживается более тесное расположение клеток небольшого размера на уровне слоя IV, воспринимающих афферентные импульсы из подкорки; в связи с этим, в отличие от полей двигательной зоны, поля лобной области характеризуются более отчетливым обособлением слоя IV. Это обстоятельство может указывать на более компактный характер афферентной сигнализации, поступающей в лобную кору от соответствующей группы ядер зрительного бугра.
Подобно тому как третичные поля задней части полушария имеют отношение к наиболее обобщенным, высоко интегрированным формам восприятия действительности, третичные поля переднелобной части полушария имеют отношение к наиболее высоко интегрированным формам целенаправленной деятельности. На всем этом мы подробно остановимся ниже (см. II, 5).
Проекционные центростремительные и центробежные связи лобной коры устанавливаются со многими подкорковыми образованиями (с определенной группой медиальных ядер зрительного бугра, подкорковыми узлами больших полушарий). Среди них особенно сильного развития достигают у человека связи с мозжечковой системой, что обусловлено прямохождением и значением мозжечковых координации для осуществления целенаправленных действий. Наряду с проекционными поля лобной области обладают обширными двусторонними ассоциационными связями с полями как прецентральной области, так и других областей коры, лежащих позади центральной борозды, а также на внутренней и нижней поверхностях полушария.
Посредством этих ассоциационных связей, а также тех связей, которые функционально объединяют разные отделы коры через подкорковые образования, обеспечивается совместная работа всех формаций коры передней части и коры задней части больших полушарий. Таким путем достигается функциональное единство всех высших психических процессов. Информация от органов чувств, поступающая в ядерные зоны анализаторов и подвергающаяся сложному анализу и синтезу в их зонах перекрытия, передается затем в двигательную и лобную области, где происходит перешифровка ее в серии организованных в пространстве и времени двигательных импульсов, осуществляемых под постоянным контролем обратной сигнализации, идущей как от самих эффекторов, так и от органов чувств, воспринимающих движение.