

Открытие генетической роли Днк
К 1944 г. О. Эйвери и его коллеги К. Маклеод и М. Маккарти открыли трансформирующую активность ДНК у пневмококков. Эти авторы продолжили работу Гриффита, описавшего феномен трансформации (передачи наследственных признаков) у бактерий. О. Эйвери, К. Маклеод, М. Маккарти показали, что при удалении белков, полисахаридов и РНК трансформация бактерий не нарушается, а при воздействии на индуцирующее вещество ферментом дезоксирибонуклеазой трансформирующая активность исчезает.
В этих экспериментах впервые была продемонстрирована генетическая роль молекулы ДНК. В 1952 г. А. Херши и М. Чейз подтвердили генетическую роль молекулы ДН К в опытах на бактериофаге Т2. Пометив его белок радиоактивной серой, а ДНК-радиоактивным фосфором,они инфицировали этим бактериальным вирусом кишечную палочку Е. coli. В потомстве фага было выявлено большое количество радиоактивного фосфора и лишь следы S. Отсюда следовало, что именно ДНК, а не белок фага проникает в бактерию, а затем после репликации передается фаговому потомству.
В 1953 г. двум американским ученым - генетику Джеймсу Уотсону и физику Френсису Крику удалось разгадать, как устроена молекула ДН К. Предложенная ими модель была основана на данных рентгено-структурного анализа ДН К, проведенного Морисом Уилкинсом и Розалинд Франклин, и учитывала правило эквивалентности Чаргаффа, согласно которому молярные отношения пуриновых и пиримидиновых азотистых оснований в ДНК близки к 1:1, т.е. содержание А = Т, a C = G. Дальнейшие исследования показали, каким образом относительно просто устроенная молекула справляется со своей генетической ролью.
Оказалось, что ДНК передает наследственную информацию в матричных процессах репликации и транскрипции. Наличие же всего четырех азотистых оснований, которые в различных сочетаниях организованы в триплетные кодирующие единицы - кодоны, не ограничивает возможности выполнения этой молекулой своих генетических функций, поскольку различная последовательность оснований и их соотношение (A+T)/(G+C) обеспечивают генетическое разнообразие видов и индивидуумов.
Уже в середине 19 века было установлено, что способность к наследованию тех или иных признаков организмов связана с материалом, содержащимся в клеточном ядре. В 1868-72 гг. швейцарский биохимик И. Ф. Мишер выделил из клеток гноя (лейкоцитов) и спермы лосося вещество, которое им было названо нуклеином, а впоследствии получило название дезоксирибонуклеиновая кислота.
В конце 19 — начале 20 вв. благодаря работам Л. Кесселя, П. Левена, Э. Фишера и др. было установлено, что молекулы ДНК представляют собой линейные полимерные цепи, состоящие из многих тысяч соединенных друг с другом мономеров — дезоксирибонуклеотидов четырех типов. Эти нуклеотиды образованы остатками пятиуглеродного сахара дезоксирибозы, фосфорной кислоты и одним из четырех азотистых оснований: пуринов — аденина и гуанина и пиримидинов — цитозина и тимина. Для обозначения оснований стали использовать начальные буквы их названий на английском или русском (в русскоязычной научной литературе) языке: соответственно A, G (Г), С (Ц) и Т.
Долгое время считалось, что ДНК содержится только в клетках животных, пока в 1930-х гг. российским биохимиком А. Н. Белозерским не было показано, что ДНК является обязательным компонентом всех живых клеток. Первые доказательства генетической роли ДНК (как вещества наследственности) были получены в 1944 группой американских ученых (О. Эйвери и др.), которые в опытах на бактериях однозначно установили, что с ее помощью наследуемый признак может быть перенесен от одной клетки к другой.
ДНК является носителем генетической информации, записанной в виде последовательности нуклеотидов с помощью генетического кода. С молекулами ДНК связаны два основополагающих свойства живых организмов — наследственность и изменчивость. В ходе процесса, называемого репликацией ДНК, образуются две копии исходной цепочки, наследуемые дочерними клетками при делении, таким образом образовавшиеся клетки оказываются генетически идентичны исходной.
Строение нуклеотида ДНК. Типы нуклеотидов
Нуклеотиды состоят из остатка фосфорной кислоты и нуклеозида, нуклеозид состоит из азотистого основания и дезоксирибозы или рибозы. Образуют полимерную цепь. Схематически это можно показать так:
 Сахар,
входящий в состав
нуклеотида,
содержит пять углеродных атомов, т. е.
представляет собой пентозу. В зависимости
от вида пентозы, присутствующей в
нуклеотиде, различают два типа нуклеиновых
кислот — рибонуклеиновые кислоты (РНК),
которые содержат рибозу, и
дезоксирибонуклеиновые кислоты (ДНК),
содержащие дезоксирибозу. В дезоксирибозе
— ОН-группа при 2-м атоме углерода
заменена на атом Н, т. е. в ней на один
атом кислорода меньше, чем в рибозе.
Сахар,
входящий в состав
нуклеотида,
содержит пять углеродных атомов, т. е.
представляет собой пентозу. В зависимости
от вида пентозы, присутствующей в
нуклеотиде, различают два типа нуклеиновых
кислот — рибонуклеиновые кислоты (РНК),
которые содержат рибозу, и
дезоксирибонуклеиновые кислоты (ДНК),
содержащие дезоксирибозу. В дезоксирибозе
— ОН-группа при 2-м атоме углерода
заменена на атом Н, т. е. в ней на один
атом кислорода меньше, чем в рибозе.
В обоих типах нуклеиновых кислот содержатся основания четырех разных видов: два из них относятся к классу пуринов и два — к классу пиримидинов. Основной характер этим соединениям придает включенный в кольцо азот. К числу пуринов относятся аденин (А) и гуанин (Г), а к числу пиримидинов — цитозин (Ц) и тимин (Т) или урацил (У) (соответственно в ДНК или РНК). Тимин химически очень близок к урацилу (он представляет собой 5-метилурацил, т. е. урацил, в котором у 5-го углеродного атома стоит метильная группа). В молекуле пуринов имеется два кольца, а в молекуле пиримидинов — одно.
Основания принято обозначать первой буквой их названия: А, Г, Т, У и Ц.
Структурная организация молекул ДНК (модель Уотсона и Крика)
ДНК – дезоксирибонуклеиновая кислота – биологическая макромолекула, носитель генетической информации во всех эукариотических клетках. Трехмерная модельпространственного строения двухцепочечной ДНК была описана в 1953 г. Дж. Уотсоном и Френсисом Криком. Согласно этой модели молекула ДНК состоит из двух полинуклеотидных цепей, которые образуют правую спираль (винтовую линию) относительно одной и той же оси. Направление цепей взаимно противоположное. Структура ДНК – полимер, структурнойединицей которого является нуклеотид. Нуклеотид состоит из: азотистого основания: пуринового – аденин (А) или гуанин (Г) или пиримидинового – цитозин (Ц) или тимин (Т); углевода дезоксирибозы (пятиуглеродное сахарное кольцо); остатка фосфорной кислоты (НРО3*).
Двойная спираль ДНК правосторонняя. 10 пар оснований составляют полный оборот 360о, следовательно, каждая пара оснований повернута на 36 о вокруг спирали относительно следующей пары. Сахарофосфатный остов располагается по периферии двойной спирали, а азотистые основания находятся внутри и их плоскости перпендикулярны оси спирали (рис. 2). Между основаниями образуются специфические водородные связи, в результате чего осуществляетсяся так называемое уотсон–криковскоеспаривание. Аденин всегда образует водородные связи с тимином, а гуанин с цитозином.
Такая закономерность называется комплементарностью. Комплементарность это определенная последовательностей оснований в противоположных цепях ДНК. Данная закономерность очень важна для репликации ДНК.

Рис. 2.
Схема структуры участка ДНК
Свойства и функции ДНК
ДНК является носителем генетической информации, записанной в виде последовательности нуклеотидов с помощью генетического кода. С молекулами ДНК связаны два основополагающих свойства живых организмов — наследственность и изменчивость. В молекуле ДНК с помощью биологического кода зашифрована последовательность аминокислот в пептидах. Важными свойствами генетического кода является его специфичность (каждый триплет способен кодировать только одну аминокислоту), универсальность (свидетельствует о единстве происхождения всего живого на Земле) и неперекрываемость кодонов при считывании. ДНК выполняет следующие функции: - хранение наследственной информации (происходит с помощью гистонов.) - передача наследственного материала (происходит путем репликации ДНК) - реализация наследственной информации в процессе синтеза белка.

Какие же из перечисленных выше структурных и функциональных особенностей молекулы ДНК позволяют ей хранить и передавать наследственную информации от клетки к клетке, от поколения к поколению, обеспечивать новые комбинации признаков у потомства?
1. Стабильность. Она обеспечивается водородными, гликозидными и фосфодиэфирными связями, а также механизмом репарации спонтанных и индуцированных повреждений;
2. Способность к репликации. Благодаря этому механизму в соматических клетках сохраняется диплоидное число хромосом. Схематично псе перечисленные особенности ДНК как генетической молекулы изображены на рисунке.
3. Наличие генетического кода. Последовательность оснований в ДНК с помощью процессов транскрипции и трансляции преобразуется в последовательность аминокислот в полипептидной цепи; 4. Способность к генетической рекомбинации. Благодаря этому механизму образуются новые сочетания сцепленных генов.
Ауторепродукция ДНК. Репликон и его функционирование
Деление клетки, как практически все остальные важнейшие клеточные процессы, начинается в ядре. Первый этап — репликация (дупликация) всех хромосом, только после этого может начаться митоз. Репликация ДНК начинается за 5-10 ч до митоза и длится 4-8 ч. Ее результатом является образование двух точных копий всей ДНК. Во время митоза каждой из дочерних хромосом достается по одной из этих копий. Между окончанием репликации и началом митоза проходит 1-2 ч. В течение этого времени в клетке происходят подготовительные процессы, которые в итоге перерастают в митоз.
Химические и физические превращения в ходе репликации ДНК. Репликация ДНК происходит почти так же, как и транскрипция РНК на матрице ДНК, за исключением нескольких важных отличий.
1. Реплицируется не одна, а обе цепи ДНК каждой хромосомы. 2. Обе цепи ДНК реплицируются полностью — от одного конца до другого, а не частично, как при транскрипции РНК. 3. В отличие от РНК-полимеразы ДНК-полимераза представляет собой комплекс основных ферментов репликации. Этот комплекс прикрепляется к ДНК и начинает двигаться вдоль нее. Другой фермент — ДНК-лигаза, который катализирует образование связей между соседними нуклеотидами, используя для этого энергию фосфатных связей.
4. Дочерние цепи ДНК начинают формироваться одновременно в сотнях участков обеих родительских цепей. Впоследствии концы отдельных сегментов вновь синтезированной ДНК «сшиваются» ферментом ДНК-лигазой. 5. Каждая вновь синтезированная цепь ДНК остается прикрепленной посредством слабых водородных связей к родительской цепи, используемой в качестве матрицы. Впоследствии обе цепи ДНК вместе скручиваются в спираль. 6. Каждая цепь ДНК имеет длину около 6 см и состоит из миллионов витков, поэтому раскрутить две цепи без специального механизма было бы невозможно. Это достигается с помощью ферментов, которые регулярно разрезают каждую спираль по всей длине, поворачивают ее фрагменты так, чтобы они могли расплестись, и затем вновь восстанавливают целостность каждой спирали. Так возникают две новые спирали.
Репарация ДНК, коррекция ДНК и мутации. Как уже упоминалось, между завершением репликации и началом митоза проходит около 1 ч. Все это время в клетке идут активные процессы репарации и коррекции ДНК. Если во время репликации к нуклеотиду материнской цепи ДНК присоединяется некомплементарный нуклеотид дочерней цепи, то с помощью ферментов он будет вырезан и заменен на комплементарный. Эти ферменты представляют собой те же самые ДНК-полимеразы и ДНК-лигазы, которые используются в процессе репликации. Этот процесс называют коррекцией ДНК.
Благодаря репарации и коррекции ДНК ошибки транскрипции, называемые мутациями, встречаются очень редко. Появление мутаций приводит к синтезу в клетке дефектных белков вместо нормальных, вследствие этого ее функции часто нарушаются, и она может даже погибнуть. Геном человека содержит не менее 30000 генов, и период между двумя поколениями составляет в среднем 30 лет, поэтому любой геном, унаследованный от родителей, должен нести не менее 10 мутаций. Однако от этих мутаций можно найти защиту. Как известно, человеческий геном представлен двойным набором хромосом, поэтому из двух аналогичных генов хотя бы один почти наверняка будет нормальным.
Спираль ДНК характеризуется рядом параметров:
- ширина спирали около 2 нм;
- шаг или полный оборот спирали составляет 3,4 нм и содержит 10 пар комплементарных нуклеотидов.
ДНК обладает уникальными свойствами: способностью к самоудвоению (репликации) и способностью к самовосстановлении (репарации).
20 белков: узнающих измененные участки ДНК и удаляющие их из цепи, восстанавливающих правильную последовательность нуклеотидов и сшивающих восстановленный фрагмент с остальной молекулой ДНК.5% всей клеточной РНК.
Репликация осуществляется под контролем ряда ферментов и протекает в несколько этапов. Она начинается в определенных точках молекулы ДНК. Специальные ферменты разрывают водородные связи между комплементарными азотистыми, и спираль раскручивается. Поинуклеотидные цепи материнской молекулы удерживаются в раскрученном состоянии и служат матрицей для синтеза новых цепей.
С помощью фермента ДНК-полимеразы из имеющихся в среде трифосфатов дезоксирибонуклеотидов (дАТФ, дГТФ, дЦТФ, дТТФ) комплементарно материнским цепям собираются дочерние цепи. Репликация осуществляется одновременно на обеих материнских цепях, но с разной скоростью и снекоторыми отличиями. На одном из цепей (лидирующей) сборка дочерней цепи идет непрерывно, на другой (отставщей) - фрагментароно. В последующем синтезированные фрагменты смешиваются с помощью фермента ДНКлигазы. В результате из одной молекулы ДНК образуется две, каждая и которых имеет материнскую и дочернюю цепи. Синтезированные молекулы являются точными копиями друг друга и исходной молекулы ДНК. Такой способ репликации ДНК называется полуконсервативным и обеспечивает точное воспроизведение в дочерних молекулах той информации, которая была записана в материнской молекуле.
Живые организмы в течение S-фазы клеточного цикла, которая предшествует делению клетки, удваивают содержание ДНК таким образом, что каждая дочерняя клетка после деления получает набор хромосом, идентичный родительской клетке. Процесс удвоения хромосом называют репликацией (редупликацией).
Хромосома содержит одну непрерывную двухцепочечную молекулу ДНК. При репликации каждая цепь родительской двухцепочеч-ной ДНК служит матрицей для синтеза новой комплементарной цепи. Вновь образованная двойная спираль имеет одну исходную (родительскую) и одну вновь синтезированную (дочернюю) цепь. Такой механизм удвоения ДНК получил название"полуконсервативная репликация" (рис. 4-14). Первичная структура дочерней цепи определяется первичной структурой родительской цепи, потому что в основе её образования лежит принцип комплементарно-сти оснований (G ≡ С и А = Т).
Ферменты и белки, участвующие в репликации, должны работать быстро и точно. Эти условия выполняются с помощью особого мультиферментного комплекса.
Репликацию можно разделить на 4 этапа: образование репликативной вилки (инициация), синтез новых цепей (элонгация), исключение праймеров, завершение синтеза двух дочерних цепей ДНК (терминация).
А. Инициация репликации
Синтез ДНК у эукариотов происходит в S-фазу клеточного цикла. Инициацию репликации регулируют специфические сигнальные белковые молекулы - факторы роста. Факторы роста связываются рецепторами мембран клеток, которые передают сигнал, побуждающий клетку к началу репликации (см. раздел 11).
Синтез новых одноцепочечных молекул ДНК может произойти только при расхождении родительских цепей. В определённом сайте (точка начала репликации) происходит локальная денатурация ДНК, цепи расходятся и образуются две репликативные вилки, движущиеся в противоположных направлениях.
В образовании репликативной вилки принимает участие ряд белков и ферментов. Так, семейство ДНК-топоизомераз (I, II и III), обладая нуклеазной активностью, участвует в регуляции суперспирализации ДНК. Например, ДНК-топоизомераза I разрывает фосфоэфирную связь в одной из цепей двойной спирали и ковалентно присоединяется к 5'-концу в точке разрыва (рис. 4-15). По окончании формирования репликативной вилки фермент ликвидирует разрыв в цепи и отделяется от ДНК.
Разрыв водородных связей в двухцепочечной молекуле ДНК осуществляет ДНК-хеликаза.Фермент ДНК-хеликаза использует энергию АТФ для расплетения двойной спирали ДНК.
В результате происходит раскручивание участка суперспирализованной молекулы ДНК. В поддержании этого участка ДНК в раскрученном состоянии участвуют SSB-белки (от англ, single strand binding proteins, т.е. белки, связывающиеся с одноцепочечными нитями ДНК). SSB-белки, не закрывая азотистых оснований, связываются с одноцепочечной ДНК по всей длине разделившихся цепей и таким образом предотвращают их комплементарное скручивание и образование "шпилек". Они обладают большим сродством к одноцепочечным участкам ДНК, независимо от первичной структуры цепей.

Рис. 4-14. Полуконсервативная репликация.
150

Рис. 4-15. Участие ДНК-топоизомеразы I в образовании репликативной вилки. 1 - фермент расщепляет одну цепь ДНК; между остатком тирозина молекулы фермента и фосфорным остатком цепи образуется ковапентная связь; 2 - происходит локальное раскручивание двойной спирали при участии ДНК-хеликазы; ДНК-топоизомераза I восстанавливает фосфоэфирную связь.
Б. Элонгация
Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами (рис. 4-16). Субстратами и источниками энергии для синтеза продукта служат 4 макроэргических соединения - дезоксирибонуклеозидтрифосфаты дАТФ, дГТФ, дЦТФ и дТТФ, для активации которых необходимы ионы магния. Нейтрализуя отрицательный заряд нуклеотидов, они повышают их реакционную способность. Ферменты проявляют каталитическую активность только в присутствии предварительно раскрученной матричной двухцепочечной ДНК. Синтез цепей ДНК происходит в направлении 5'→3' растущей цепи, т.е. очередной нуклеотид присоединяется к свободному 3'-ОН-концу предшествующего нуклеотид-ного остатка. Синтезируемая цепь всегда антипараллельна матричной цепи. В ходе репликации образуются 2 дочерние цепи, представляющие собой копии матричных цепей.
В синтезе эукариотических ДНК принимают участие 5 ДНК-полимераз (α, β, γ, δ, ε). ДНК-полимеразы различают по числу субъединиц, молекулярной массе, ассоциации с разными вспомогательными белками, ускоряющими процесс биосинтеза ДНК, и функциональному назначению. ДНК-полимеразы α (альфа), β (бета), δ (дельта), ε (эпсилон) участвуют в синтезе ДНК в ядре клеток, ДНК-полимераза γ (гамма) - в репликации митохондриальной ДНК.
ДНК-полимеразы β, δ, ε не могут инициировать образование дочерних цепей, так как не имеют сродства к одиночной нити ДНК. Инициирует репликацию ДНК-полимераза α, которая комплементарна определённому сайту одноцепо-чечной ДНК. Присоединяясь к нему, ДНК-полимераза а синтезирует небольшой фрагмент РНК - праймер, состоящий из 8-10 рибонуклеотидов. ДНК-полимераза а состоит из четырёх субъединиц. Каждая из субъединиц фермента выполняет определённую функцию: "узнавание" сайта репликации, синтез праймера (8-10 рибо-нуклеотидов), синтез фрагмента цепи ДНК, около 50 дезоксирибонуклеотидов. Таким образом, ДНК-полимераза α синтезирует олигонуклеотид, содержащий примерно 60 нуклеотидных остатков; первые 8-10 представлены рибонуклеотида-ми (праймер), а остальные - дезоксирибонуклеотидами.
ДНК-полимераза δ
Олигонуклеотид, синтезированный ДНК-полимеразой α и образующий небольшой двухцепочечный
151

Рис. 4-16. Репликация.
фрагмент с матрицей, позволяет присоединиться ДНК-полимеразе δ и продолжить синтез новой цепи в направлении от 5'- к 3'-концу по ходу раскручивания репликативной вилки.
ДНК-полимераза δ последовательно наращивает цепь, шаг за шагом присоединяя к ней соответствующие дезоксинуклеотиды. Выбор ДНК-полимеразой δ очередного нуклеотида определяется матрицей. Включение дезоксирибонуклеозидмонофосфатов в растущую цепь ДНК сопровождается гидролизом макроэргических связей соответствующих нуклеозидтрифосфатов и отщеплением пирофосфата (Н4Р2О7). Энергия макроэргических связей расходуется на образование 3',5'-фосфодиэфирной связи между последним нуклеотидом растущей цепи ДНК и присоединяемым нуклеотидом. Включение нуклеотида в синтезируемую цепь ДНК невозможно без предварительного связывания азотистого основания водородными связями с комплементарным нуклеотидом матричной цепи. ДНК-полимеразы (α, β, γ, δ, ε) могут синтезировать нуклеотидную цепь только в направлении 5'→3', матричная цепь всегда считывается в направлении 3'→5'.
В каждой репликативной вилке идёт одновременно синтез двух новых (дочерних) цепей. Направление синтеза цепи ДНК совпадает с направлением движения репликативной вилки лишь для одной из вновь синтезируемых цепей (лидирующая цепь). На второй матричной цепи синтез дочерней ДНК осуществляется двумя ферментами: ДНК-полимеразой α и ДНК-полимеразой ε в направлении 5'→3', но против движения репликативной вилки. Поэтому вторая цепь синтезируется прерывисто, короткими фрагментами, которые называют "фрагменты Оказаки" (по имени открывшего их исследователя). Дочерняя цепь ДНК, синтез которой происходит фрагментами, называют отстающей цепью. Каждый фрагмент Оказаки, примерно 100 нуклеотидных остатков, содержит праймер. Праймеры удаляет ДНК-полимераза β, постепенно
152
отщепляя с 3'-конца фрагмента по одному ри-бонуклеотиду. К ОН-группе на 3'-конце предыдущего фрагмента ДНК-полимераза β присоединяет дезоксирибонуклеотиды в количестве, равном вырезанному праймеру и таким образом заполняет брешь, возникающую при удалении рибонуклеотидов.
Фермент ДНК-лигаза катализирует образование фосфодиэфирной связи между 3'-ОН-группой дезоксирибозы одного фрагмента цепи ДНК и 5'-фосфатом следующего фрагмента. Реакция протекает с затратой энергии АТФ. Таким образом, из множества фрагментов Оказаки образуется непрерывная цепь ДНК.
В. Ориджины репликации
ДНК хромосомы человека содержит примерно 150 млн пар нуклеотидов. Репликация такой большой молекулы со скоростью 50 нуклеотидов в минуту шла бы примерно 800 ч. Поэтому инициация синтеза ДНК происходит в нескольких сайтах хромосомы, которые называют сайтами инициации репликации, или ориджинами (от англ. origin - происхождение) репликации (рис. 4-17). Термин "сайт" используют для обозначения любого участка генома. Ориджины репликации имеют определённую нуклеотидную последовательность. Последовательность ДНК, ограниченную двумя ориджинами репликации, называют единицей репликации, или репликоном. На ориджинах при участии ДНК-топоизомеразы I инициируется двунаправленная репликация. Образуются две репликативные вилки, перемещающиеся в противоположных направлениях до тех пор, пока не встретятся со следующим репликоном, т.е. репликация прекращается, когда встречаются две репликативные вилки.
Метилирование ДНК
После завершения репликации происходит метилирование нуклеотидных остатков вновь образованных цепей ДНК. Метальные группы присоединяются ко всем остаткам аденина в последовательности -GATC-, при этом образуется N6-метиладенин, а также возможны метилирование цитозина в последовательности -GC-и образование N5-метилцитозина. Количество метилированных оснований равно примерно 1-8%. Модификация происходит при участии ферментов, использующих в качестве источника метальных групп S-аденозилметионин (SAM) (см. раздел 9). Присоединение метальных групп к остаткам аденина и цитозина не нарушает комплементарности цепей (рис. 4-18).
Наличие метальных групп в цепях ДНК необходимо для формирования структуры хромосом, а также для регуляции транскрипции генов. В течение непродолжительного времени в молекуле ДНК последовательности -GATC-метилированы по аденину только в матричной, но не в новой цепи. Это различие используется ферментами репарации для исправления ошибок, которые могут возникать при репликации.
Г. Строение 3'- и 5'-концов цепей ДНК. Теломерная ДНК
На каждом конце хромосомы присутствует специфическая нуклеотидная последовательность. Она представлена многочисленными повторами (сотни или даже тысячи раз) олигонуклеотидов

Рис. 4-17. Образование двух репликативных вилок, перемещающихся в противоположных направлениях от ориджина.
153

Рис. 4-18. Метилирование остатков аденина в последовательности -GATC-. В течение нескольких минут после репликации, пока не произошло метилирование, новая цепь ДНК отличается от матричной цепи.
-GGGTTA-, называемых теломерной последовательностью, или просто теломерной ДНК. Наличие теломер необходимо для завершения репликации концевых информативных последовательностей хромосом, т.е. для сохранения генетической информации.
После завершения репликации хромосомы 5'-концы дочерних цепей ДНК недостроены, так как после удаления праймеров эти фрагменты оказываются недореплицированными. Это происходит потому, что ДНК-полимераза β, отвечающая за заполнение бреши, образованной после удаления праймера, не может вести синтез цепи ДНК от 3'- к 5'-концу (рис. 4-19, А). Таким образом, в ходе каждого цикла репликации 5'-концы синтезированных цепей укорачиваются. Но такие потери не представляют опасности для генетической информации хромосом, потому что укорочение ДНК идёт за счёт теломер. Во время следующего цикла репликации
154
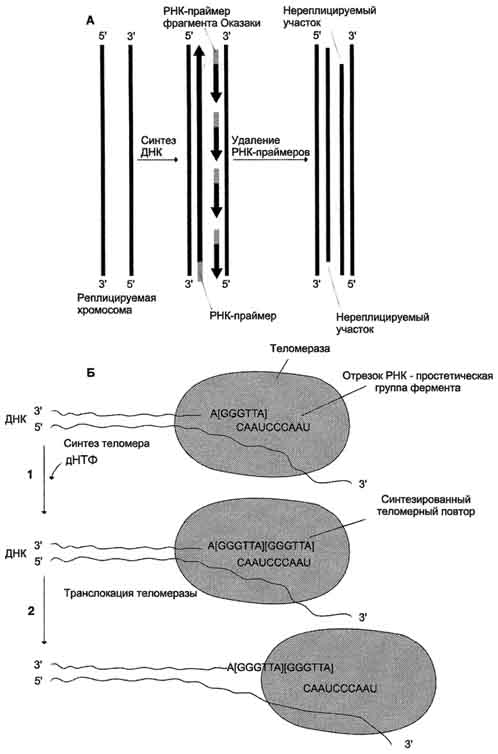
Рис. 4-19. Синтез теломерной ДНК. А - на рисунке показано укорочение вновь синтезированных цепей ДНК после удаления праймеров; Б - в состав теломеразы входит короткая молекула РНК, содержащая в активном центре последовательность нук-леотидов, комплементарную теломерному повтору; 1 - фермент прикрепляется за счёт взаимодействия РНК с существующей теломерой и добавляет последовательно по одному нуклеотиду фрагмент -GGGTTA-. Матрицей служит простетическая группа теломеразы - фрагмент РНК; 2 - фермент перемещается по нити ДНК таким образом, что РНК-матрица в составе теломеразы постоянно комплементарно связана с концом вновь синтезированного теломерного повтора. Заново синтезированная тело-мерная ДНК служит матрицей для удлинения второй цепи ДНК, но уже в ходе следующего цикла клеточного деления. Теломер-ный повтор на рисунке взят в квадратные скобки -[GGGTTA]-.
155
5'-концы цепей ДНК опять остаются недостроенными. Таким образом, с каждым клеточным делением ДНК хромосом будут последовательно укорачиваться. Укорочение теломер в большинстве клеток по мере их старения - важный фактор, определяющий продолжительность жизни организма.
Однако в эмбриональных и других быстро-делящихся клетках потери концов хромосом недопустимы, потому что укорочение ДНК будет происходить очень быстро. В эукариотических клетках имеется фермент теломераза (нуклео-тидилтрансфераза), обеспечивающий восстановление недореплицированных 5'-концов. К особенностям этого фермента относят присутствие в качестве простетической группы РНК. Фрагмент РНК в активном центре теломеразы служит матрицей при синтезе теломер-ных повторов хромосом.
С помощью РНК фермент комплементарно прикрепляется к 3'-концу недостроенной дочерней цепи ДНК. Теломераза по принципу комплементарности последовательно удлиняет 3'-конец цепи ДНК на один гексануклеотид -GGGTTA-. Синтез всегда идёт от 5'- к 3'-концу. Затем теломераза смещается по цепи ДНК на один теломер и начинает синтез нового фрагмента -GGGTTA- (рис. 4-19, Б).
В большинстве соматических клеток теломераза неактивна, так как соматическая клетка имеет длину теломерной ДНК, достаточную для времени жизни клетки и её потомства. Однако небольшую активность теломеразы обнаруживают в клетках с высокой скоростью обновления, таких как лимфоциты, стволовые клетки костного мозга, клетки эпителия, эпидермиса кожи и др.
Генетический код, его свойства
Генетический код – система записи генетической информации в ДНК (РНК) в виде определенной последовательности нуклеотидов. Последовательность нуклеотидов определяет последовательность включения АК в синтезируемый белок. 3 нуклеотида – триплет – кодон – кодируют 1 АК. Совокупность триплетов и составляет генетический код.
Свойства генетического кода:
1) триплетность
2) вырожденность (избыточность – 1 АК кодируется несколькими триплетами)
3) специфичность (1 кодон кодирует только 1 АК)
4) однонаправленность (от 5’ к 3’ концу)
5) неперекрываемость (один нуклеотид входит в состав только одного кодона)
6) универсальность (у всех живых организмов одинаковые АК кодируются одинаковыми кодонами)
7) отсутствие знаков препинания внутри гена.
1. Код триплетен. В состав РНК входят 4 нуклеотида: А, Г, Ц, У. Если бы мы пытались обозначить одну аминокислоту одним нуклеотидом, то 16 из 20 аминокислот остались бы не зашифрованы. Двухбуквенный код позволил бы зашифровать 16 аминокислот (из четырех нуклеотидов можно составить 16 различных комбинаций, в каждой из которых имеется два нуклеотида). Природа создала трехбуквенный, или триплетный, код. Это означает, что каждая из 20 аминокислот зашифрована последовательностью трех нуклеотидов, называемых триплетом или кодоном. Из 4 нуклеотидов можно создать 64 различные комбинации по 3 нуклеотида в каждой (4*4*4=64). Этого с избытком хватает для кодирования 20 аминокислот и, казалось бы, 44 кодона являются лишними. Однако это не так.
2. Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном (от двух до шести). Исключение составляют аминокислоты метионин и триптофан, каждая из которых кодируется только одним триплетом. (Это видно из таблицы генетического кода .) Тот факт, что метионин кодируется одним триплетом АУТ, имеет особый смысл, который вам станет понятен позже ( 16 ).
3. Код однозначен. Каждый кодон шифрует только одну аминокислоту. У всех здоровых людей в гене, несущем информацию о бета-цепи гемоглобина , триплет ГАА или ГАГ, I стоящий на шестом месте, кодирует глутаминовую кислоту. У больных серповидноклеточной анемией второй нуклеотид в этом триплете заменен на У. Как видно из таблицы, триплеты ГУА или ГУГ, которые в этом случае образуются, кодируют аминокислоту валин. К чему приводит такая замена, вы уже знаете из раздела о ДНК .
4. Между генами имеются "знаки препинания". В печатном тексте в конце каждой фразы стоит точка. Несколько связанных по смыслу фраз составляют абзац. На языке генетической информации таким абзацем являются оперон и комплементарная ему и-РНК. Каждый ген в опероне кодирует одну полипептидную цепочку - фразу. Так как в ряде случаев по матрице и-РНК последовательно создается несколько разных полипептидных цепей, они должны быть отделены друг от друга. Для этого в генетическом коде существуют три специальные триплета - УАА, УАГ, УГА, каждый из которых обозначает прекрдщение синтеза одной полипептидной цепи. Таким образом, эти триплеты выполняют функцию знаков препинания. Они находятся в конце каждого гена. Внутри гена нет "знаков препинания". Поскольку генетический код подобен языку, разберем это свойство на примере такой составленной из триплетов фразы: жил был кот тих был сер мил мне тот кот. Смысл написанного понятен, несмотря на отсутствие "знаков препинания. Если же мы уберем в первом слове одну букву (один нуклеотид в гене), но читать будем также тройками букв, то получится бессмыслица: илб ылк отт ихб ылс ерм илм нет отк от Нарушение смысла возникает и при выпадении одного или двух нуклеотидов из гена. Белок, который будет считываться с такого испорченного гена, не будет иметь ничего общего с тем белком, который кодировался нормальным геном.
6. Код универсален. Генетический код един для всех живущих на Земле существ. У бактерий и грибов, пшеницы и хлопка, рыб и червей, лягушки и человека одни и те же триплеты кодируют одни и те же аминокислот
Понятие о гене. Свойства гена
Ген (др.-греч. γένος — род) — структурная и функциональная единица наследственности живых организмов.
Ген представляет собой последовательность ДНК, задающую последовательность определённого полипептида либо функциональной РНК. Гены (точнее, аллели генов) определяют наследственные признаки организмов, передающиеся от родителей потомству при размножении. При этом некоторые органеллы (митохондрии, пластиды) имеют собственную, определяющую их признаки, ДНК, не входящую в геном организма.
Свойства гена
дискретность — несмешиваемость генов;
стабильность — способность сохранять структуру;
лабильность — способность многократно мутировать;
множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм;
аллельность — в генотипе диплоидных организмов только две формы гена;
специфичность — каждый ген кодирует свой признак;
плейотропия — множественный эффект гена;
экспрессивность — степень выраженности гена в признаке;
пенетрантность — частота проявления гена в фенотипе;
амплификация — увеличение количества копий гена.
Особенности организации генома эукариот
У эукариотических организмов механизм регуляции транскрипции гораздо более сложен. В результате клонирования и секвенирования генов эукариот обнаружены специфические последовательности, принимающие участие в транскрипции и трансляции. Для эукариотической клетки характерно: 1. Наличие интронов и экзонов в молекуле ДНК. 2. Созревание и-РНК - вырезание интронов и сшивка экзонов. 3. Наличие регуляторных элементов, регулирующих транскрипцию, таких как: а) промоторы - 3 вида, на каждый из которых садится специфическая полимераза. Pol I реплицирует рибосомные гены, Pol II - структурные гены белков, Pol III - гены, кодирующие небольшие РНК. Промотор Pol I и Pol II находятся перед участком инициации транскрипции, промотор Pol III - в рамках структурного гена; б) модуляторы - последовательности ДНК, усиливающие уровень транскрипции; в) усилители - последовательности, усиливающие уровень транскрипции и действующие независимо от своего положения относительно кодирующей части гена и состояния начальной точки синтеза РНК; г) терминаторы - специфические последовательности, прекращающие и трансляцию, и транскрипцию. Эти последовательности по своей первичной структуре и расположению относительно инициирующего кодона отличаются от прокариотических, и бактериальная РНК-полимераза их не "узнает". Таким образом, для экспрессии эукариотических генов в клетках прокариот нужно, чтобы гены находились под контролем прокариотических регуляторных элементов. Это обстоятельство необходимо учитывать при конструировании векторов для экспрессии.
Геном эукариот
В ходе эволюции у эукариот сформировался геном, который существенно отличается от генома прокариот. Геном высших организмов, в первую очередь, характеризуется таким свойством как избыточность. Содержание ДНК у эукариот в расчете на 1 клетку в среднем на 2–3 порядка выше, чем у прокариот. Хотя такой размер генома теоретически позволяет кодировать свыше 106 различных белков, оценки числа функционирующих генов у эукариот колеблются в интервале от 104 до 27105.
Возможно, существуют "молчащие" гены, необходимые клетке для реализации редко встречающихся программ. Однако это не является единственной причиной большого количества ДНК в клетках эукариот. Повышение количества ДНК у эукариот связано, прежде всего, с усложнением регуляторных процессов в клетках. Это усложнение выражается в том, что размеры единиц транскрипции у эукариот, как правило, на порядок превышают размеры зрелых молекул иРНК, а также в том, что значительную часть генома у эукариот составляют последовательности, служащие исключительно для целей регуляции.
Однако общее количество ДНК в клетке нельзя рассматривать как простой показатель функциональной и структурной сложности организма. Например, у некоторых рыб количество ДНК на ядро в 10 раз больше, чем у человека. Содержание ДНК может сильно варьировать даже у отдельных организмов одного рода. Все это говорит о реальной избыточности ДНК в клетках высших организмов.
Другая структурная особенность генома эукариот – это блочный характер организации. Блочность строения проявляется на разных уровнях. Это и длинные блоки с разным содержанием, и многочисленные короткие участки, сильно обогащенные определенными (пуриновыми или пиримидиновыми) нуклеотидами, и гомополимерные последовательности. Блочность строения проявляется и в структуре индивидуальных генов, которые часто состоят из отделенных друг от друга зон (экзонов и интронов). Наличие генов с интронами характерно для эукариот, тогда как в геноме прокариот интронов не обнаружено. Геному эукариот свойственна также способность к перестройкам.
Компактизация ДНК важна для организмов по двум причинам:
1) позволяет упорядоченно расположить очень длинные молекулы ДНК в небольшом объеме без потери доступности для считывания информации;
2) это один из способов дифференциальной регуляции активности разных участков генома, т.к. активность генов зависит от характера упаковки ДНК. У прокариот компактизация ДНК достигается сверхспирализацией и петельной организацией.
У эукариот прослеживается иерархия из 4 уровней упаковки ДНК от нуклеосомной нити до структуры хромосомы. В обоих случаях организация структуры генетического материала осуществляется при помощи специальных ДНК-связывающих белков.
Геном эукариот устроен намного сложнее, чем прокариотический геном, что связано, прежде всего, с повышением уровня сложности регуляции генетических процессов, а не за счет увеличения количества структурных белков и ферментов в клетке. Блочный характер строения генома эукариот является еще одной реализацией принципа комбинаторного увеличения разнообразия: например, за счет объединения "мини"-генов (экзонов) в один "супер"-ген могут возникать функционально новые белки. В эволюции генома, вероятно, активную роль играют мобильные генетические элементы.
Классы нуклеотидных последовательностей в ДНК эукариот, характеристика, свойства, биологическое значение
Нуклеотидная последовательность, генетическая последовательность — порядок следования нуклеотидных остатков в нуклеиновых кислотах. Определяется при помощи секвенирования.
Секвенирование биополимеров (белков и нуклеиновых кислот — ДНК и РНК) — определение их первичной аминокислотной или нуклеотидной последовательности (отлат. sequentum — последовательность). В результате секвенирования получают формальное описание первичной структуры линейной макромолекулы в виде последовательности мономеров в текстовом виде.
Уровни организации генома эукариот
Химический и структурный состав хромосом