

Просты́е белки́ — белки, которые построены из остатков α-аминокислот и при гидролизе распадаются только на аминокислоты.
Простые белки по растворимости в воде и солевых растворах условно подразделяются на несколько групп: протамины, гистоны, альбумины, глобулины, проламины, глютелины.
Альбумины и глобулины широко распространены в органах и тканях животных. В плазме крови человека в норме содержится 7% белков, представленных преимущественно альбуминами и глобулинами. Альбумины и глобулины – глобулярные белки, различающиеся по растворимости.
Необходимо отметить, что само определение «альбумины» и «глобулины» основано на их растворимости в дистиллированной воде и полунасыщенном растворе (NH4)2SO4. Однако, как показывают данные табл. 1.6, глобулины растворимы только в разбавленных солевых растворах.

Различную растворимость альбуминов и глобулинов сыворотки крови раньше широко использовали в клинической практике для их фракционирования и количественного определения.
В настоящее время качественный состав и содержание сывороточных белков определяют с помощью электрофореза на бумаге и в полиакрил-амидном геле в небольшом количестве сыворотки крови. Альбумины и глобулины отличаются друг от друга также по молекулярной массе – соответственно 40000–70000 и 150000 и более.
Глобулины (globulinum; лат. globulus, уменьшительное от globus шар)
— общее название белков, растворимых в слабых растворах нейтральных солей, кислот и щелочей, как правило, нерастворимых в дистиллированной воде и выпадающих в осадок при 50% насыщении растворов сульфатом аммония; глобулины составляют около 40% всех белков сыворотки крови человека.
Гамма-глобулин - любой белокБЕЛОК (protein) - органическое соединение, в состав которого входят углерод, водород, кислород и азот (..., присутствующий в плазме крови, который можно идентифицировать по характерной скорости его движения в электрическом полеПОЛЕ - 1) в социологии - изучаемая среда, аудитория, которую опрашивают в ходе полевых исследований (в... Фракция γ-глобулинов является наиболее гетерогенной. Известно множество антител, различающихся первичной структурой. Электрофоретически они открываются главным образом в γ-глобулиновой и частично в β2-глобулиновой фракциях.
К неоднородной группе гамма-глобулинов относятся белки с самой низкой электрофоретической подвижностью. К ним относится большинство защитных веществ крови, многие из которых обладают ферментативной активностью. Так как потребности в белках, выполняющих такие специальные функции, бывают различны, размеры и состав фракции гамма-глобулинов может значительно изменяться. Почти при всех заболеваниях, особенно воспалительных , содержание гамма-глобулинов в плазме крови повышается. В то же время общее количество белков в плазме обычно остается примерно одинаковым, так как повышение содержания гамма-глобулинов сопровождается уменьшением фракции альбумина, в результате снижается так называемый альбумин-глобулиновый коэффициент. Почти все гаммаглобулиныГАММАГЛОБУЛИН (gamma globulin) - любой белок, присутствующий в плазме крови, который можно идентифицировать по ... являются иммуноглобулинами
Иммуноглобулины.
История открытия.
Иммуноглобулины представляют собой белки человека (животных), которые обычно обладают свойствами антител, т.е. специфической способностью соединяться с антигеном, который стимулирует их образование. Иммуноглобулины присутствуют в крови, цереброспинальной жидкости, лимфоузлах, селезенке, слюне и других тканях, а так же в виде рецепторов на поверхностных мембранах клеток. Синтезируются они в В-лимфоцитах, содержат углеводные группировки и могут рассматриваться как гликопротеины. По электрофоретической подвижности иммуноглобулины относятся в основном к гамма-глобулинам и бета2-глобулинам. Биологическая роль иммуноглобулинов в организме связана с участием в процессах иммунитета. Их защитная функция обусловлена способностью специфически взаимодействовать с антигенами. До середины 50-х годов прошлого столетия о структурной организации иммуноглобулинов ничего не знали. Первый шаг в этом направлении сделал английский иммунохимик Р.Портер в 1959 г. Он показал, что при обработке очищенных иммуноглобулинов протеолитическими ферментами образуются три фрагмента, два из которых взаимодействуют с антигеном (патогеном) и потому названы антигенсвязывающими (Fab), и один, неспособный к такому взаимодействию (Fc).
Но это ничего не говорило о причинах их специфичности по отношению к разным антигенам. Для получения информации о молекулярных основах вариабельности нужно было значительное количество полностью идентичных белков. Сывороточные иммуноглобулины, в массе образующиеся после иммунизации, не давали такой возможности, поскольку они - производные нескольких клеточных клонов, каждый из которых продуцирует белки только одного класса и только одной, свойственной ему, специфичности. Иначе говоря, иммуноглобулины, выделяемые от иммунизированных животных, есть смесь молекул с разной специфичностью и разной принадлежностью к тому или иному классу.
Необходима была экспериментальная модель, позволяющая работать с одним клоном, образующим иммуноглобулины только одной специфичности и только одного класса. Природа предоставила такую возможность - злокачественно трансформированные плазматические клетки больных миеломой. В настоящее время существует большой набор клонированных плазмоцитов человека и животных, продуцирующих соответствующие миеломные белки.
Строение.(см. пример: строение IgG)
Изучение аминокислотной последовательности иммуноглобулинов выявило принципиальные особенности в их строении.
Молекулы иммуноглобулинов симметричны. Они построены из "легких" (около 220 аминокислотных остатков, молекулярная масса 25000 для IgG) и "тяжелых" (450-600 аминокислотных остатков, молекулярная масса для IgG- 50000) полипептидных цепей (соотв. L- (light) и Н-цепи (heavy)), скрепленных дисульфидными связями и нековалентными взаимодействиями. В антителах человека обнаружено два вида легких цепей ( и ) и пять видов тяжелых цепей ( и ), отличающихся аминокислотной последовательностью. Тяжелые цепи, характерные для каждого из классов и подклассов иммуноглобулинов, содержат по одному или более олигосахаридному фрагменту (степень гликозилированности отражается на биологических свойствах иммуноглобулинов).
Р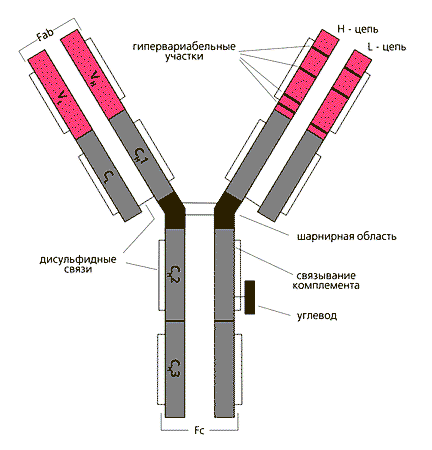 ис.1.
Строение
иммуноглобулина G.
ис.1.
Строение
иммуноглобулина G.
Две тяжелые (Н) цепи с мол. весом 50 кД и две легкие (L) с мол. весом 25 кД объединены в единую молекулу с помощью ковалентных дисульфидных связей. Каждая цепь содержит вариабельную область (VL и VH для L- и H-цепей соответственно) и константную (С), подразделяющуюся у Н-цепей на гомологичные участки (домены): CH1, CH2, CН3. L-цепь имеет один константный участок - CL. От взаимодействия VH- и VL-областей зависит специфичность иммуноглобулинов как антител. В аминокислотной последовательности V-доменов имеются гипервариабельные участки, характеризующиеся частой заменой аминокислот от белка к белку, и более консервативные. Между СН1 и СН2 доменами Н-цепи находится шарнирная область, обеспечивающая подвижность антигенсвязывающего Fab-фрагмента. СН2-домен служит местом присоединения углеводов и связывания комплемента. СН3-домен взаимодействует с Fc-рецептором (не способным связывать антиген) на поверхности клеток, принимающих участие в иммунологических реакциях.
Лёгкие цепи различных видов отличаются друг от друга С - концевой последовательностью аминокислот. С - концевая половина полипептидной цепи имеет постоянную аминокислотную последовательность, а ее N - концевая часть - вариабельную. Каждый постоянный (Сl) и вариабельный (Vl) участок легкой цепи включает 107-110 аминокислотных остатков. Тяжелые цепи построены их четырех участков - VH, C1H, C2H, C3H. Вариабельные участки их состоят приблизительно из 110-114 аминокислотных остатков, постоянные - 330.
В вариабельной части полипептидных цепей находятся определенные, так называемые "гипервариабельные участки", с наибольшим числом аминокислотных замен. В легких цепях они расположены между 24-34; 52-55; 89-97 аминокислотными остатками. Гипервариабельные участки тяжелых цепей занимают аналогичные положения, но точная локализация их пока не установлена.
Строение постоянных областей тяжелых цепей определяет эффекторные функции молекул к поверхностям макрофагов, В-лимфоцитов, тучных клеток, а также проникновение через плацентарную мембрану.
Гибкость молекул иммуноглобулинов, обеспечивающая приспособляемость к различным конфигурациям молекул антигена, обусловливается также наличием особого "шарнирного участка" в середине тяжелых цепей, содержащего много остатков аминокислоты пролина и препятствующего образованию вторичной структуры. Шарнирная область чувствительна к протеолитическим ферментам. При расщеплении ими (например, папаином) иммуноглобулин распадается на два идентичных Fab-фрагмента и один Fc-фрагмент.
В настоящее время на основании изучения первичной структуры полипептидных цепей выдвинута так называемая "доменная" гипотеза строения иммуноглобулинов, согласно которой молекулу иммуноглобулинов можно разбить на участки с относительно независимыми конфигурациями в виде глобул. Каждый домен состоит приблизительно из 100-110 аминокислотных остатков и имеет одну дисудьфидную связь, которая связывает участки цепей, образуя петлю из 60 аминокислотных остатков.
Молекулы иммуноглобулинов связанные с поверхностью лимфоцитов, имеют дополнительные гидрофобные "хвосты" на С-концах тяжелых цепей, которые встроены в мембраны клеток. Пептидные цепи иммуноглобулинов и ряда белков клеточных мембран (антигены гистосовместимости, рецепторы для антигенов Т-лимфоцитов) по своей первичной структуре сходны между собой, что указывает на общее эволюционное происхождение всех этих белков.
Связывание с антигенами.
На N-концах тяжелых и легких цепей расположены те самые вариабильные области, которые в сочетании и образуют антигенсвязывающую структуру - паратоп в составе Fab-фрагмента. Три или четыре домена со стороны С-концов тяжелых цепей составляют константную часть молекулы - Fc-фрагмент (не связывают антиген). Fc-Фрагмент состоит из четырех или шести доменов двух тяжелых цепей и определяет такие свойства иммуноглобулинов, как связывание ими комплемента, возможность проникать через плаценту, присоединяться к клеткам и фиксироваться в коже. Поскольку в состав молекулы иммуноглобулина входят две легкие и две тяжелые цепи, они формируют два паратопа (области, контактирующие с антигеном) в составе двух Fab-фрагментов, т.е. антитело бивалентно: может соединиться с двумя идентичными антигенными эпитопами. Этому способствует наличие шарнирной области между первым и вторым доменами константного фрагмента тяжелых цепей, благодаря которой обеспечивается возможность пространственной ориентации Fab-фрагментов для связывания с антигенными эпитопами. Комплекс с антигеном образуется в результате нековалентных взаимодействий, характер которых может варьировать в зависимости от специфичности антитела- связи могут быть ионными, вандерваальсовыми, водородными, при помощи солевых мостиков и гидрофобных взаимодействий.. Сила связывания с антигеном увеличивается на несколько порядков, если молекула антитела реагирует сразу двумя (или более) областями связывания с несколькими детерминантами одной молекулы антигена.
Каждая индивидуальная клетка вырабатывает антитела только одной
специфичности по правилу "одна клетка " одно антитело" (Петров , 1987). Это
означает, что в клетке активно функционируют только один вариант гена VH,
один - гена СH и по одному соответствующему гену одной из легких цепей. Все
остальные структурные гены выключены. В каждой отдельно взятой антитело-
образующей клетке из всего множества структурных генов иммуноглобулинов
функционирует их минимальное количество, необходимое для синтеза антител
одной специфичности и одного типа. Таким образом, в основе многообразия
специфичности антител лежит функционирование в лимфоидной системе большого
количества клеток и их потомков - клонов клеток - продуцентов одного вида
антител. Следовательно, количество специфичностей антител соответствует
количеству клонов клеток-антителопродуцентов, различающихся генами,
функционирующими в них.
Генетика.
Особенностью этих полипептидных цепей является отсутствие единого гена, кодирующего структуру всей полипептидной цепи. Всякий раз сборка такого гена происходит из отдельных сегментов. Этим обеспечивается бесконечное разнообразие структур молекул антител, способных распознать любую существующую в природе структуру антигена. Иными словами, набор (репертуар) специфических участков связывания в популяции иммуноглобулинов организма столь широк, что на любой попадающий в организм антигенный эпитоп (участок связывания) обязательно найдется строго комплементарный паратоп в составе антиген-связывающего фрагмента (Fab - фрагмента) какого-то иммуноглобулина. Поэтому говорят, что легкие и тяжелые пептидные цепи каждого класса иммуноглобулинов построены из двух основных областей - вариабельной и постоянной.
Все их антигенные детерминанты кодируются тремя несцепленными группами аутосомных генов. Одна группа кодирует тяжелую цепь
того или иного класса, другая - легкую k- типа, третья – легкую -типа. Так как полипептидные цепи состоят из двух различных участков - вариабельного (V) и постоянного (С), каждая из трех групп генов включает набор генов вариабельной - V и постоянной областей -С-гены. Таким образом, синтез каждой полипептидной цепи молекулы иммуноглобулина контролируется двумя структурными генами, а не одним, как при синтезе других белков. Один ген кодирует вариабельную область цепи, другой- постоянную. Причем существуют многие гены для вариабельных
областей полипептидной цепи, что связано с многообразием специфических
активных центров.