

Жимулёв Курс лекций
.pdf
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Глава 7. Структура гена |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
Íà |
|
Ðèñ. |
7.57. |
суммированы |
|
последовательности |
могут |
иметь |
||||||||||||||||||||||
возможные механизмы образования |
регуляторные последствия. |
|
|
|
||||||||||||||||||||||||||
многих типов молекул РНК, считываемых |
|
|
|
На Рис. 7.58. представлена схема |
|
|||||||||||||||||||||||||
с одного гена: за счет изменения |
альтернативного сплайсинга в гене BR-C |
|||||||||||||||||||||||||||||
инициации и терминации транскрипции, |
|
у дрозофилы. Весь ген занимает около 120 |
||||||||||||||||||||||||||||
а также сплайсинга. Использование |
т.п.н., в нем выделяют 10 экзонов. |
|||||||||||||||||||||||||||||
альтернативных промоторов |
может |
|
Обнаружено более 15 типов различных |
|
||||||||||||||||||||||||||
изменять |
5' |
конец, |
использование |
ìÐÍÊ. |
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
альтернативных терминаторов может |
|
|
Другой |
òèï |
альтернативного |
|||||||||||||||||||||||||
изменять 3' конец транскрипта. Эти |
сплайсинга, |
когда |
îäèí |
ãåí |
|
|||||||||||||||||||||||||
изменения могут организоваться только |
функционирует фактически как два (Рис. |
|
||||||||||||||||||||||||||||
инициирующими или терминальными |
7.59.). Показаны два транскрипта: Е74А и |
|
||||||||||||||||||||||||||||
последовательностями мРНК. Иногда |
Е74В, синтезирующиеся одновременно с |
|||||||||||||||||||||||||||||
изменения |
â |
нетранслируемой |
|
5’ |
двух разных промоторов. |
|
|
|
|
|||||||||||||||||||||
лидерной последовательности или 3’ |
|
|
Альтернативный сплайсинг играет |
|||||||||||||||||||||||||||
нетранскрибируемой |
|
трэйлерной |
огромную |
|
ðîëü |
â |
генетическом |
|
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
определении пола (см. Рис. 13.6.). |
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
Промоторы Экзон 1 |
Экзон 2 |
Экзон 3 |
Экзон 4 |
Экзон 5 Терминаторы |
|
|
|
|||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
2 |
3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
2 |
3 |
|
|
|
|
||||
|
|
|
|
|
|
Использование альтернативных промоторов |
|
|
|
|
|
|
||||||||||||||||||
|
|
|
|
|
|
|
|
|
Экзоны 1 |
2 |
|
3 |
4 |
|
|
5 |
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
2 |
3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
Промоторы |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
Использование альтернативных терминаторов |
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
1 |
|
2 |
|
3 |
4 |
|
5 |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Терминаторы
1 2
Альтернативный сплайсинг
1 |
2 |
3 |
5 |
|
|
|
|
|
|
|
|
|
|
|
1 |
4 |
5 |
2 |
|
|
|
|||
Терминатор
2
Рис. 7.57. Схема возможных вариантов альтернативного сплайсинга (Из: Lewin, 1994, p. 688)
7-62
Глава 7. Структура гена
à
á
â
100 |
120 |
140 |
160 |
|
|
|
|
|
|
180 |
|
|
|
|
200 |
|
220 ò.ï.í. |
|||||||||
экзоны |
1 |
|
|
2 |
|
3 4 5 |
6 |
|
7 |
|
8 |
9 |
|
10 |
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
z1 |
|
z2 |
z3 |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
коровый |
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
экзон |
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|




 A n
A n
An
An
An
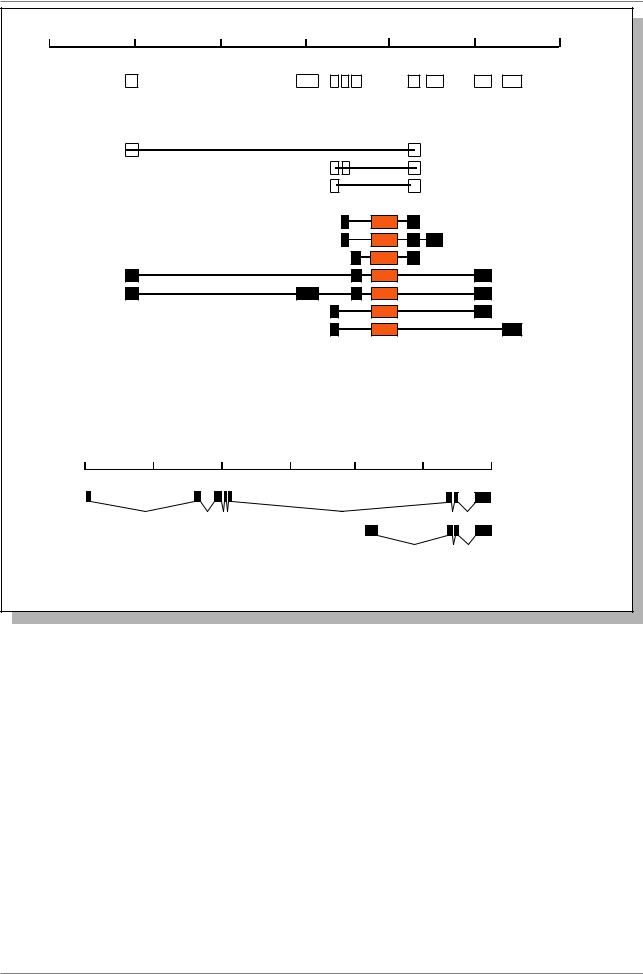
Рис. 7.58. Организация транскрипции гена BR-C у D. melanogaster (Из: DiBello et al., 1991, в кн. Жимул¸в, 1994, стр. 284); а - физическая карта гена (от 100 до 220 т.п.н.); б - расположение экзонов (1-10) на физической карте; в - различные транскрипты, возникающие за счет альтернативного сплайсинга
5’ 0 |
10 |
20 |
30 |
40 |
50 |
60 ò.ï.í. 3’ |
E74A
E74B
Рис. 7.59. Структура гена Е74, расположенного в пуфе 74EF (Из: Karim, Thummel, 1991, в кн. Жимулев, 1994, стр. 293)
Во время созревания макронуклеуса |
содержит 14 экзонов, расположенных в |
||||
у инфузорий |
происходит |
удаление |
следующем порядке: 1, 3, 5, 7, 9, 11, 2, 4, 6, |
||
внутригенных последовательностей без |
8, 10, 12, 13, 14, а нормальный по |
||||
синтеза РНК. Более того, происходит |
нумерации порядок экзонов образуется |
||||
престановка |
экзонов. |
Òàê, |
â |
в созревшем макронуклеусе (Рис. 7.60.). |
|
микронуклеарной ДНК экзоны гена |
7.9.3. Локализация генов в |
||||
актина I у Oxytricha nova расположены в |
|||||
последовательности: 3, 4, 6, 5, 7, 9, 2, 1, 8. |
интронах |
|
|||
После удаления “интронов” экзоны |
В интронах некоторых генов |
||||
занимают положение в ряду с 1 по 9 и |
располагаются другие гены (Рис. 7.61 - |
||||
только такая их последовательность да¸т |
7.63.). |
|
|||
функциональный белок (См. детали: |
Удивительным по |
сложности |
|||
Prescott, 1992). |
|
|
|
организации и величине |
интронов |
Ещ¸ более впечатляет реорганизация |
является ген dnc у D.melanogaster. Ген |
||||
гена αТВР, который в микронуклеусе |
занимает минимум 130 т.п.н. (от -90 до |
||||
7-63

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Глава 7. Структура гена |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
3 |
5 |
7 911 |
2 |
4 |
6 |
8 |
10 12 |
|
13 |
14 |
|
|
|
|
|
|||||||
|
|
|
|
IES 1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
IES 2 |
IES 3 |
IES 4 IES 5 IES 6 |
|
IES 7 |
IES 8 IES 9 |
|
IES 10 IES 11 IES 12 |
|
IES 13 |
||||||||||
MDS 14
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
MDS 13 |
3’ TAS |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
IES 13 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
IES 7 |
|
|
MDS 4 |
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
MDS 3 |
|
IES 2 |
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
MDS 2 |
|
|
|
|
|
|
IES 8 |
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
IES 1 |
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
IES 12 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
MDS 1 |
|
MDS 5 IES 3 MDS 6 |
|
|
|
|
|
|
|||||
|
|
|
|
|
|
MDS 12 |
|
IES 6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5’ TAS |
MDS 7 |
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
MDS 11 |
|
|
|
IES 9 |
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
IES 4 |
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
IES 11 |
|
IES 5 MDS 9 |
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
MDS 10 |
|
|
IES 10 |
|
|
|
MDS 8 |
IES 1 - IES 11 |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
(369 ï.í.) |
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
IES 12 (37 ï.í.) |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
IES 13 (58 ï.í.) |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
1 |
|
|
2 |
3 |
5 |
6 |
8 |
9 |
11 |
13 |
|
|
14 |
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
4 |
|
7 |
|
10 |
12 |
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
13 |
|
|
14 |
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
1 |
|
|
2 |
3 |
5 |
6 |
8 |
9 |
11 |
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
4 |
|
7 |
|
10 |
12 |
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
кодирующий сегмент (1539 п.н.) |
|
|
|
|
|
|
|
|
|
|||||||
теломера ATG |
|
|
|
|
|
|
|
|
|
|
TGA |
теломера |
||||||||||||||
Рис. 7.26. Схема “созревания” гена αТВР у Oxitricha nova.
а - микронуклеарный вариант гена, содержащего 14 экзоноподобных элементов (последовательностей, предназначенных для макронуклеуса - ППМ) и 13 интроноподобных (внутренних элиминируемых последовательностей - ВЭП).
б -Возможная картина образования ДНК-петель для считывани я последовательных ППМ; в - последовательность ППМ после созревания макронуклеус а;
г - зрелый ген с добавленными теломерами
+40 т.п.н.) и содержит 13 экзонов. Между первым экзоном и экзоном, расположенным еще более дистально (около 40 т.п.н.), расположено несколько генов из семейства Pig/Sgs. Между вторым и третьим экзонами (около 70 т.п.н.) располагается еще 4 гена (Рис. 7.63.).
7.9.4. Использование промоторов генов теплового шока
Организацию и экспрессию структурной части генов можно изучать, объединяя их в конструкцию с известным промотором, и трансформируя
7-64
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Глава 7. Структура гена |
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ген куколочного |
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
кутикулярного белка |
||||||||
47 ò.ï.í. |
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
0.9 ò.ï.í. |
|
|
|
||||||||||||||||
3’ ïîëè-À |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5’ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
3’ ïîëè-À 17 ò.ï.í. |
|
|
|
|
|
|
|
|
|
|||||||||
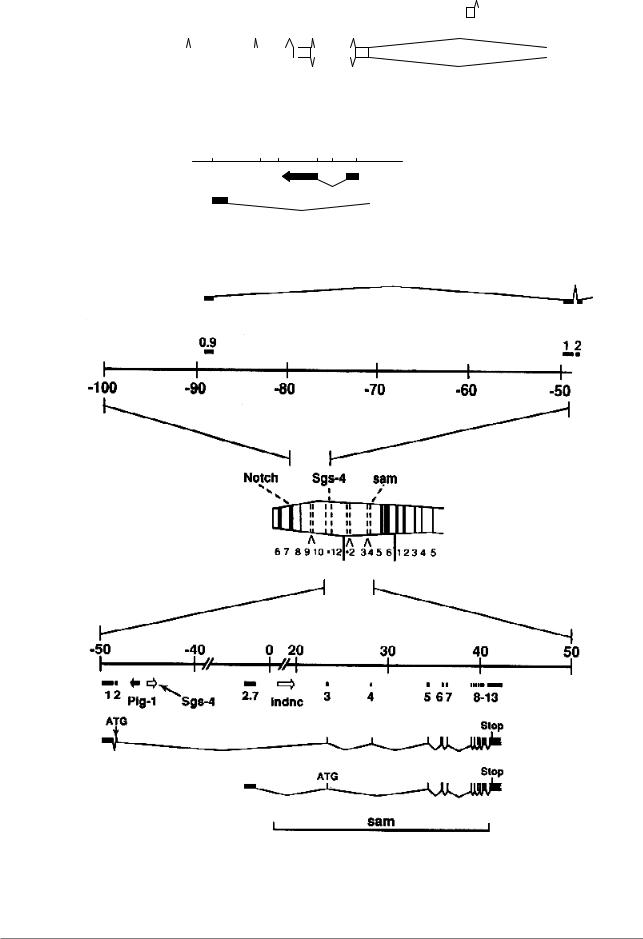
Рис. 7.61. Экзон-интронная карта гена Gart и расположенного в его интроне гена куколочного кутикулярного белка дрозофилы (Из: Жимулев, 1994, стр. 147)
R R R B S R
sina
 Rh4
Rh4
Рис. 7.62. Расположение гена sina в интроне гена Rh4 дрозофилы. Вверху - рестрикционная карта района (Из: Carthew, Rubin, 1990, в кн. Жимулев, 1994, стр. 148)
à
á
â
ã
â
á
à
Рис. 7.63. Структура гена dnc у D. melanogaster (Из: Davis, Dauwalder, 1990, в кн. Жимулев, 1994, стр. 149); а - карта интронов и экзонов гена dunce; б - карта других генов, расположенных в интронах гена dunce; в - физическая карта ДНК в районе хромосомы 3С; г - участок хромосомы 3С6 - 3Е5
7-65

Глава 7. Структура гена
дрозофил, мутантных по этому гену. Для этих целей используют промоторы генов теплового шока. В нормальном развитии эти помоторы не функционирует, но активируется при резком повышении температуры (370С на 30 мин) (см. раздел 12.).
Ген под действием шока начинает экспрессироваться, о чем можно судить по “исправлению” мутантного фенотипа, появлению нормального белка или РНК (см. детали: Lawrence, 1992, p. 55).
7.9.5. Участки, терминирующие транскрипцию
Терминация транскрипции прокариотических генов обусловлена элементами, называемые терминаторами. Одним из важных белков, вовлеченных в терминацию транскрипции некоторых генов у E. coli, является белок ρ. Терминаторы в таких генах называются ρ-зависимыми (или терминаторы II типа). Во многих других терминаторах сердцевинная РНК-полимераза сама
может проводить события терминации. Терминаторы такого типа называются ρ- независимыми терминаторами (терминаторы I типа). ρ-независимые терминаторы состоят из последовательностей, расположенных за 16-20 п.н. от точки терминации и представляющих собой инвертированный повтор (Рис. 7.64.).
Эта последовательность замыкается нитью из 4-8 AT-последовательностью, на которой синтезируются цепь из U- последовательностей.
ρ-зависимые терминаторы не имеют AT нити и во многих случаях не формируют шпилечных структур. ρ- фактор - это протеин с двумя доменами: один связывается с РНК, другой домен - с АТФ.
Три ключевые события происходят на терминаторах обоих типов:
1.останавливается синтез РНК,
2.цепь РНК освобождается от ДНК,
3.РНК-полимераза освобождается от ДНК.
Матрица |
5’ |
CCCAGCCCGCCTAATGAGCGGGCTTTTTTTTGAACAAAA 3’ |
|||||||||||||||||||||
(ÄÍÊ) |
3’ |
GGGTCGGGCGGATTACTCGCCCGAAAAAAAACTTGTTTT 5’ |
|||||||||||||||||||||
Транскрипт |
5’ CCCAGCCCGCCUAAUGAGCGGGCUUUUUUUU-OH 3’ |
||||||||||||||||||||||
(ÐÍÊ) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
A |
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
U |
|||||||||||||||||
|
|
|
|
|
|
|
|
A |
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
U |
|
|
|
|
|
|
|
|
|
|
G |
|
||||||
|
|
|
Мутации |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
C |
|
|
|
A |
|
Мутации |
||||||||||||||
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
A, U |
|
|
|
|
|
C-G |
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
Транскрипт |
|
A |
|
|
|
|
|
G-C |
|
|
|
|
A |
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
U |
|
|
|
|
|
C-G |
|
|
|
|
|
A, U, C |
|
||||||||
|
образует |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
C-G |
|
|
|
|
|
A, U |
|
|||||||
|
терминирующую |
|
|
|
|
|
|
||||||||||||||||
|
|
C-G |
|
|
|
|
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
шпильку |
G-C |
|
|
|
|
|
|
|
|
||
|
5’ CCCA-UUUUUUUU-OH 3’ |
||||
|
|
|
|
|
|
|
|
|
Делеция |
||
|
G |
||||
|
|
|
|
||
Рис. 7.64. Последовательность нуклеотидов в ρ-независимом терминаторе транскрипции и структура терминируемой РНК. Мутации в зат емненных участках полностью предотвращают терминацию (Из: Russell, 1998, p. 388)
7-66

Глава 7. Структура гена
7.9.6. Гомология генов
(не расписано)
О гомологии свидетельствует закон гомологических рядов Вавилова. Обладают гомологией:
1.Гены домашнего хозяйства (рРНК, гистонов)
2.Гены, выполняющие одинаковые функции
3.Домены в составе генов (цинковые пальцы, гомео-домен)
4.Гены теплового шока (книга по тепловому шоку Ashburner, Shlesinger)
7.9.7. Псевдогены
(не расписано)
Иногда в геномах встречаются т.н. псевдогены. Они имеют все необходимые черты генов, т.е. полный набор экзонов, характерных для этого гена, поли-А/Т хвост и короткие прямые повторы (как у мобильных элементов) в ДНК мишени.
Псевдоген начинается с 5' точки, эквивалентной 5' точке мРНК, а заканчивается трактом поли-А нуклеотидов, что вероятно имеет происхождение от поли-А конца в молекуле мРНК. (Из: Lewin, 1994, p.10511055, см. также главу 24).
У дрозофилы псевдогены встречаются редко, чаще это гены транспортных или малых ядерных РНК (тРНК, snRNA). Еще один известный пример - псевдоген личиночного кутикулярного белка и псевдоген Adh у D. mulleri, они сохраняют свои интроны. Пример процессированного псевдогена (т.е. без интронов) описан для гена Adh у D. teissieri и D. yakuba.
Литература к разделу 7.9.
Ashburner M. Drosophila. A laboratory handbook. Cold Spring Harbor. Cold SpringHarborLaboratoryPress,1-1331, 1989
Lawrence P.A. The making of fly. Oxford, Blackwell Scientific Publications, 1- 228, 1992.
Lewin B. Genes V. Oxford University Press. Oxford, New York, Tokyo, 1- 1272, 1994.
Prescott D.M. The unusual organization and processing of genomic DNA in hypotrichous ciliates. TIG 8, N12, 439-445, 1992.
Prescott D.M. The DNA of ciliated protozoa. Microbiol. Rev. 58, N2, 233-267. 1994.
Russell, P.J. Genetics Fifth edition. Addison Wesley Longman, Ins, Menlo Park, California, p. 379-417, 1998.
7-67

Глава 9. Организация и функционирование хромосом
Глава 9. Организация и функционирование хромосом
9.1. Введение |
2 |
9.2.1. Хромосомы вирусов, клеточных органелл и прокариот |
2 |
9.2.2. Геном дрожжей |
4 |
9.3. Митотические хромосомы |
5 |
9.3.1. Идентификация хромосом |
5 |
9.3.2. Кариотип и идиограмма |
6 |
9.3.3. Дифференциальные окраски хромосом |
8 |
9.3.4. “Правило Меллера” и синтения |
9 |
9.4. Эу- и гетерохроматин в митотических хромосомах |
11 |
9.4.1. Компактизация хроматина |
12 |
9.4.2. Дифференциальная окрашиваемость |
14 |
9.4.3. Конъюгация гетерохроматиновых районов |
15 |
9.4.4. Контакты гетерохроматина с ядерной оболочкой |
16 |
9.4.5. Гетерохроматин и хромосомные перестройки |
16 |
9.4.6. Поздняя репликация |
17 |
9.4.7. Варьирование количества гетерохроматина |
18 |
9.4.8. Формирование гетерохроматиновых |
|
районов хромосом в онтогенезе |
18 |
9.4.9. Повторенные оследовательности |
19 |
9.4.10. Генетическое содержание гетерохроматиновых |
|
районов хромосом |
22 |
9.4.11. Заключение |
29 |
9.5. Теломеры и теломерный гетерохроматин |
31 |
9.5.1. Концепция теломеры |
31 |
9.5.2. Строение теломер |
34 |
9.6. Диминуция хроматина и хромосом |
43 |
9.6.1. Диминуция хроматина у аскарид |
43 |
9.6.2. Диминуция хроматина у циклопов |
44 |
9.6.3. Элиминация хроматина у инфузорий |
46 |
9.6.4. Элиминация хромосом у двукрылых насекомых |
47 |
9.6.5. Физиологическое значение диминуции |
|
хроматина и хромосом |
48 |
9.7. Строение центромеры |
50 |
9.8. В-хромосомы |
53 |
9.9. Генетическая инактивация хромосомы у D. miranda |
54 |
9.10. Факультативный и конститутивный |
|
гетерохроматин |
54 |
9.11. Гетерохроматин и клетки зародышевого пути |
54 |
9-1

|
|
|
|
Глава 9. Организация и функционирование хромосом |
||||||
|
|
|
|
|
|
|||||
9. Строение и |
|
|
|
А. Вейсман |
предположил, что |
|||||
функционирование хромосом |
наследственность |
сосредоточена в |
||||||||
|
|
|
|
|
хромосомах, а доказали это Т.Х. Морган, |
|||||
9.1. Введение |
|
|
|
К. Бриджес, Г. Меллер и А. Стертевант, |
||||||
Хромосомы - это нуклеопротеиновые |
завершившие к середине 1930-х годов |
|||||||||
тела, в которых хранится, передается |
разработку |
хромосомной |
теории |
|||||||
потомству и реализуется наследственная |
наследственности. |
|
|
|
||||||
информация. |
|
|
|
|
Внастоящеевремянаиболееизвестны |
|||||
|
|
|
|
три типа хромосом: |
|
|
|
|||
По иронии судьбы сначала были |
|
|
|
|||||||
открыты ядерные структуры, которые в |
а. У прокариот в нуклеоиде и в клеточных |
|||||||||
течение многих последующих лет никто не |
органеллах у эукариот. |
|
|
|||||||
б. Хромосомы из делящихся клеток |
||||||||||
считал хромосомами. В 1881 году Э. |
||||||||||
Бальбианиописалвклеткахслюнныхжелез |
эукариот. |
|
|
|
|
|||||
в. Интерфазные хромосомы эукариот. |
||||||||||
хирономуса |
поперечно-исчерченные |
|||||||||
ленты. Их |
назвали |
“структурами |
Литература к разделу 9.1. |
|
||||||
Бальбиани”. Только в 1912 году чешский |
Босток К., Самнер Э. Хромосома |
|||||||||
ученый Ф. Рамбоусек предположил, что |
||||||||||
это специализированные хромосомы. А |
эукариотической клетки. Москва, |
|||||||||
Мир, 1-598, 1981 (перевод с англ. |
||||||||||
окончательно это название утвердилось в |
||||||||||
издания 1978 года). |
|
|
||||||||
1930-1935 ãã. (D. Kostoff, T. Painter, H. |
|
|
||||||||
Прокофьева-Бельговская А.А. Строение и |
||||||||||
Muller). |
|
|
|
|
функция хромосом в кн. Руководство |
|||||
Хромосомы, как “окрашивающиеся |
||||||||||
по цитологии, т.2, Москва-Ленинград, |
||||||||||
тела” были открыты в митотически и |
||||||||||
Наука, стр. 280-329, 1966. |
|
|||||||||
мейотически |
делящихся |
клетках |
|
|||||||
Прокофьева-Бельговская |
À.À. |
|||||||||
классиками цитологии Флеммингом и |
||||||||||
Страсбургером (W. Flemming, 1882; E. |
Гетерохроматические |
районы |
||||||||
хромосом.Москва,Наука,1-431,1986. |
||||||||||
Strasburger, 1884). Свое название |
||||||||||
|
|
|
|
|
||||||
хромосомы |
получили благодаря |
9.2.1. Хромосомы вирусов, |
||||||||
способности интенсивно окрашиваться |
клеточных органелл и |
|
||||||||
основными красителями. |
|
|
прокариот |
|
|
|
||||
Сам термин “хромосома” предложил |
|
|
|
|||||||
У ДНК - содержащих вирусов, |
||||||||||
â 1888 ãîäó W. Waldeyer. |
|
|
||||||||
|
|
бактерий, сине-зеленых водорослей, а |
||||||||
Огромное |
значение |
факта |
||||||||
такжевсамореплицирующихсяорганеллах |
||||||||||
продольного |
расщепления |
каждой |
клеток |
эукариот |
(пластидах, |
|||||
хромосомы - образование хроматиды - в |
митохондриях |
è |
центриолях) |
|||||||
процессе деления клетки отметил в тот же |
хромосома представляет собой голую |
|||||||||
период В. Ру (W. Roux, 1883). С |
двуспиральную молекулу ДНК. |
|
||||||||
удивительной интуицией он указал, что |
У большинства форм эта молекула |
|||||||||
подобный способ деления хромосом |
||||||||||
образует кольцо, которое закручено в |
||||||||||
свидетельствует о присутствии в них |
шпильку, |
и хромосома |
имеет |
|||||||
жизненно важных для клетки элементов и |
суперспирализованный вид (Рис. 9.1.). |
|||||||||
о расположении |
ýòèõ |
элементов в |
|
|
|
|
|
|||
линейном порядке.
9-2
Глава 9. Организация и функционирование хромосом
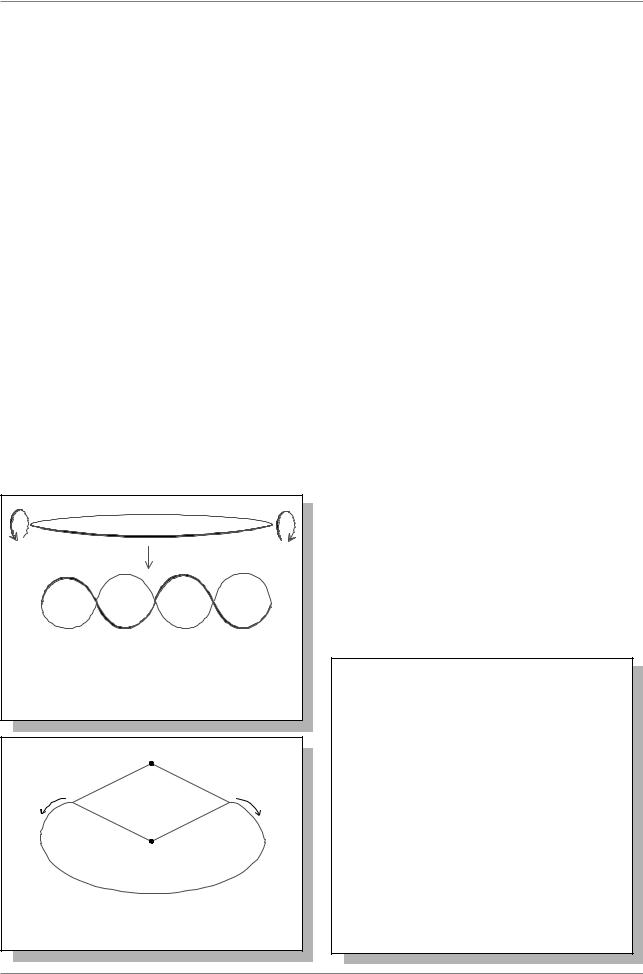
Репликация этих хромосом начинается с единственной определенной точки (точки инициации репликации) и прогрессирует, пока не закончится репликация всей хромосомы: таким образом, хромосома представляет собой единицу репликации-репликон. В большинстве случаев, а у бактерий - всегда, репликация хромосомы идет одновременно в обе стороны от точки инициации репликации (т.и.р.) (Рис. 9.2.).
Репликация хромосом вирусов и прокариот протекает быстро, со скоростью порядка 30 микронов в минуту. У прокариот хромосома прикреплена одной точкой к клеточной мембране, и после репликации расхождение дочерних хромосом осуществляется раздвиганием этих точек вследствие роста мембраны.
Ðèñ. |
9.1. |
Кольцевая |
è |
||
суперспирализованная |
форма |
||||
молекулы ДНК у бактерий |
|
|
|||
|
|
ò.è.ð. |
|
|
|
Репликация |
|
|
Репликация |
||
|
|
ò.è.ð. |
|
|
|
Ðèñ. |
9.2. |
Схема |
репликации |
||
кольцевой ДНК |
|
|
|
||
Длина молекул ДНК, служащих хромосомами у вирусов, прокариот и клеточных органелл, варьирует (Табл. 9.1.).
В результате полного секвенирования геномов некоторых бактерий длины кольцевых молекул ДНК и числа генов определены по данным на 1 сентября 1998 года: у Bacillus subtilis длина ДНК составляет 4214,814 т.п.н. и геном содержит около 4100-4220 генов, у E. coli – 4639,221 т.п.н. и, соответственно, примерно 4290 генов (Рис. 9.3.).
У бактерий геном организован в некое тело или тела, которые выглядят довольно компактными и занимают около трети обьема клетки. Эти тела называют нуклеоидами. ДНК в бактериальном нуклеоиде находится в ассоциации с ДНКсвязывающимися белками:
1.Белок HU конденсирует ДНК, возможно заворачивая ее в бусоподобные структуры. Он стимулирует репликацию ДНК.
2.Белок H1 связывается с ДНК, взаимодействуя прежде всего с теми последовательностями, которые изогнуты. Функции белка неизвестны.
Табл. 9.1. Размеры кольцевой молекулы ДНК - хромосомы у вирусов, прокариот и в клеточных органеллах эукариот (Из: Алиханян и др. 1985, стр. )
|
|
Длина |
|
Кольцевая |
|
|
|
молекула ДНК |
|
|
ïàðû |
|
микрометры |
|
|
|
|
нуклеотидов |
|
|
|
|
|
|
|
|
|
Мелкие вирусы, |
0,4-1,0 |
|
1200-3000 |
центриоли |
|
||
|
|
|
|
|
|
|
|
Другие вирусы, |
|
|
|
пластиды, |
5-100 |
|
15000-300000 |
митохондрии |
|
|
|
|
|
|
|
Бактерии |
1000-2000 |
|
3000000-6000000 |
|
|
|
|
9-3

Глава 9. Организация и функционирование хромосом
Рис. 9.3. Сопоставление генетической карты (снаружи круга) и длины кольцевой молекулы ДНК (в т.п.н.) у E. coli (данные из Internet на 1 сентября 1998 года )
3.Белок P был секвенирован и по последовательности аминокислот напоминает протамины, которые связываются с ДНК в сперматозоидах некоторых видов. Предполагается, что P - это ДНК-связывающийся белок, однако функции его не известны. ДНК в нуклеоиде составляет около 80% (для сравнения у эукариот - только 50%). Она свернута в петли, примерно по 40
т.п.н. в каждой петле (Рис. 9.4.). Основания петель защищены с
помощью какого-то неизвестного механизма, что предотвращает передачу
Рис. 9.4. Петлевая организация ДНК у бактерий
вращения ДНК с одной петли на другую. В геноме примерно 100 таких петель - или доменов. (см. детали: Lewin, 1994, pp. 772776).
Литература к разделу 9.2.
Алиханян С.И., Акифьев А.П., Чернин Л.С. Общая генетика. Москва, Высшая школа, 1-446, 1988.
Lewin B. Genes V, Oxford, New York, Tokyo, Oxford University Press, 1- 1272, 1994.
9.2.2. Геном дрожжей
Хромосомы этих низших эукариот не видны под микроскопом, однако по генетическим данным выделяют 16 групп комплементации. Геном дрожжей
Saccharomyces cerevisiae к настоящему времени полностью секвенирован. Общая длина ДНК в геноме составляет 13390 т.п.н., варьирование хромосом по длине показано на Рис. 9.5.
В геноме выявлено около 6085 генов.
9-4