Жимулёв Курс лекций
.pdf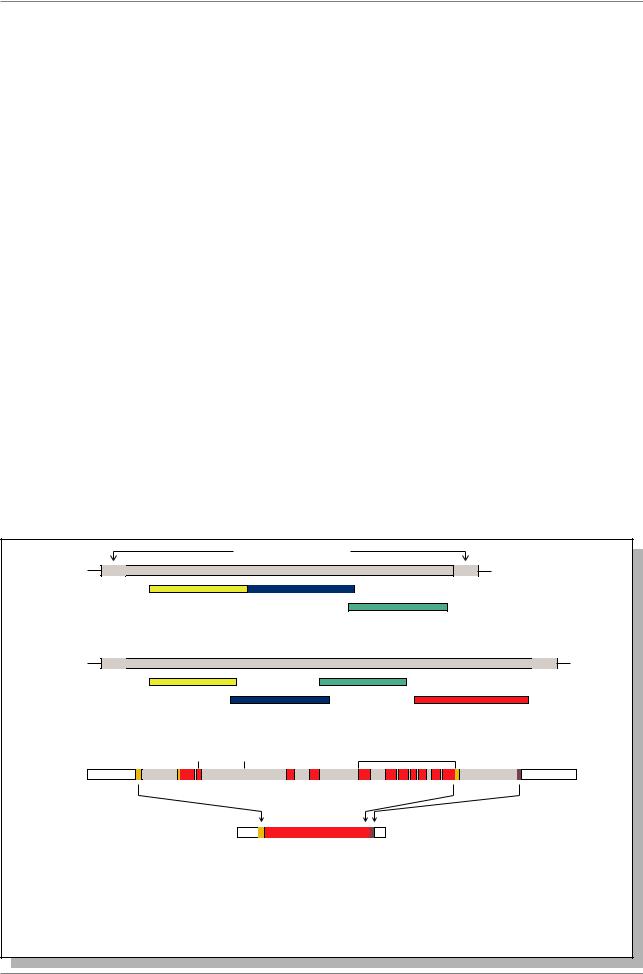


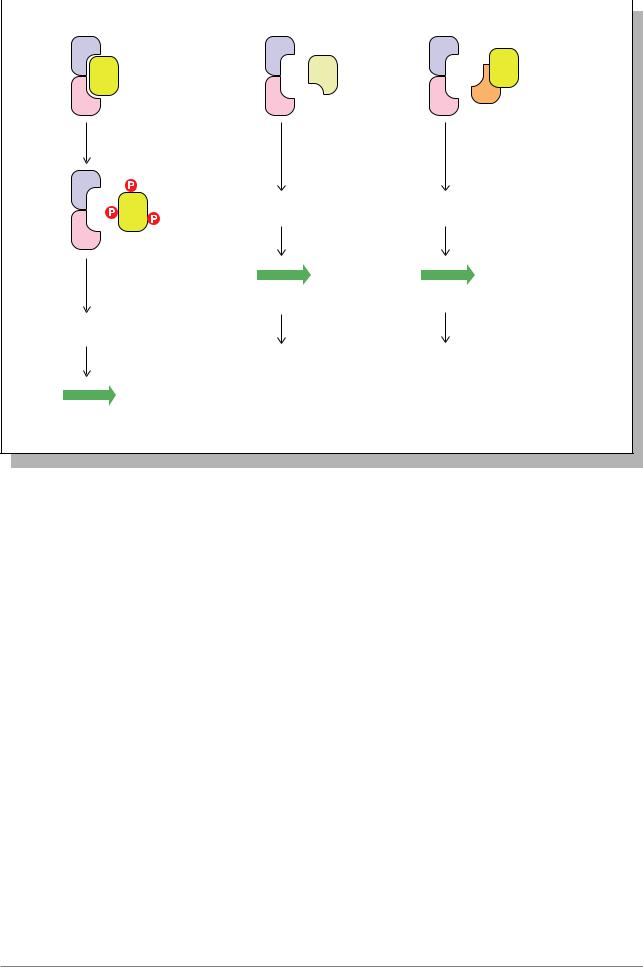
 - активация экспрессии гена
- активация экспрессии гена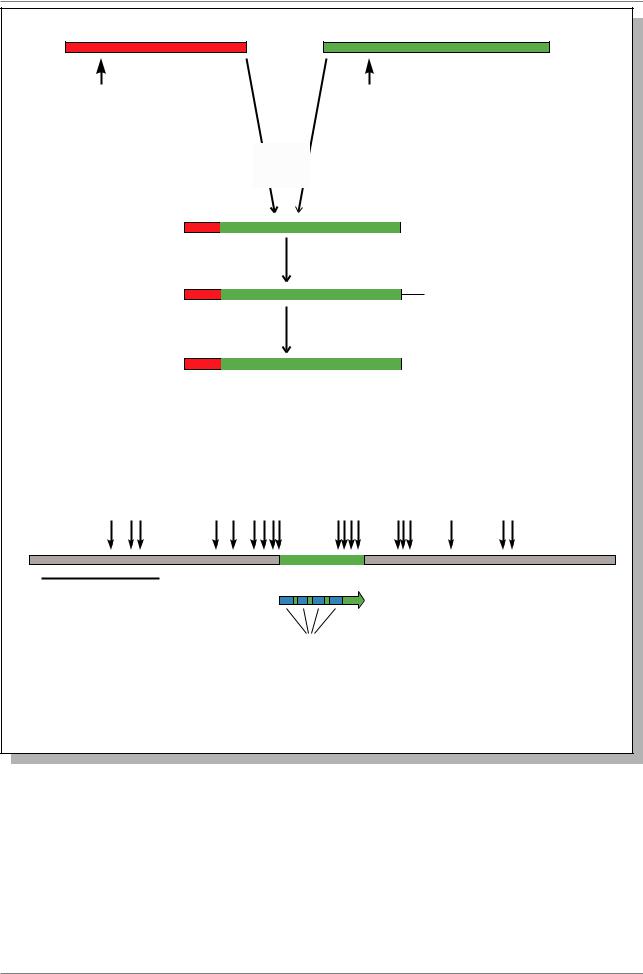
|
|
|
|
|
Глава 20. Основы онкогенетики |
||
Ãåí brc в хромосоме 22 |
|
Ãåí abl в хромосоме 9 |
|
|
|||
5’ |
|
3’ |
5’ |
|
|
3’ |
|
Точка |
|
|
|
Точка |
|
|
|
разрыва |
|
|
|
разрыва |
|
|
|
|
|
Филадельфийская |
|
|
|
||
|
|
транслокация |
|
|
|
||
|
5’ |
|
|
3’ |
ãåí bcr/abl |
|
|
|
|
|
|
|
|
|
|
|
|
|
Транскрипция |
|
|
|
|
|
5’ |
|
|
3’ |
ïîëè-A ìÐÍÊ bcr/abl |
|
|
|
|
|
|
|
|
||
|
|
|
Трансляция |
|
|
|
|
|
|
|
|
|
белок Bcr/Abl |
|
|
Рис. 20.6. Превращение протоонкогена abl в онкоген у пациентов с хронической |
|||||||
миелогенной лейкемией. В результате транслокации 22 и 9 хромосомы |
|||||||
(Филадельфийская хромосома) ген bcr соединяется с геном abl тирозиновой |
|||||||
протеинкиназы, в результате чего этот ген функционирует не по своему |
|||||||
расписанию (Из: Alberts et al., 1994, p. 1278) |
|
|
|
||||
|
|
|
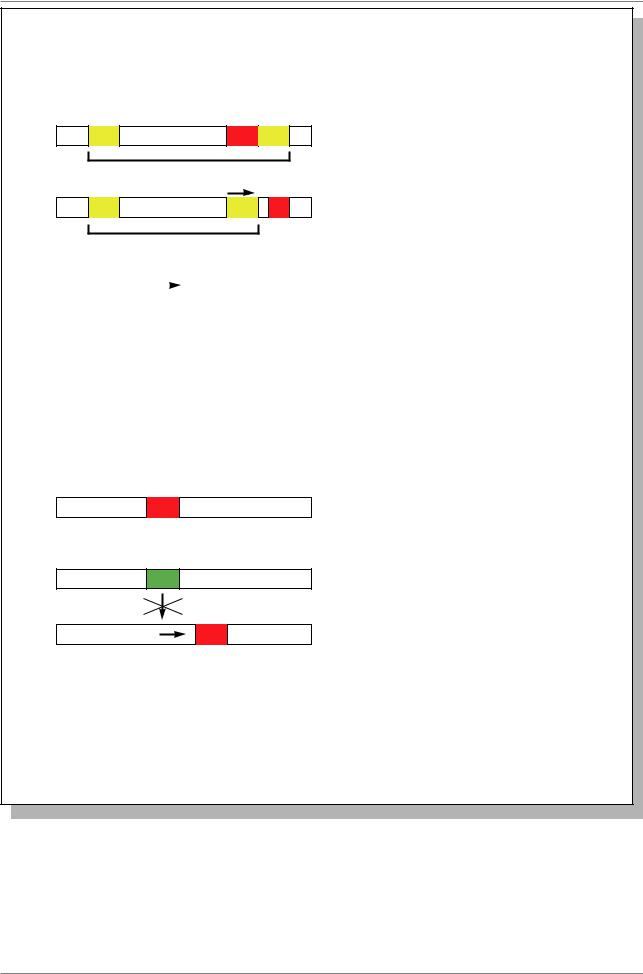
Ãåí Wnt-1 |
|
|
|
|
5’ |
|
|
|
|
|
|
3’ |
5000 ï.í. |
|
|
ÐÍÊ |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Экзоны |
|
|
|
|
Рис. 20.7. Локализация инсерций (показаны стрелками) вируса рака молочных |
|||||||
желез мыши (MMTV), активирующих транскрипцию гена Wnt-1. Инсерция |
|||||||
влияет на экспрессию гена, даже если она на расстоянии до 10 т.п.н. (Из: Alberts |
|||||||
et al., 1994, p. 1277) |
|
|
|
|
|
|
|
сетчатки глаза. Известны две формы |
Предположили, что у особей с |
||||||
заболевания - наследственное и наследственной формой утрачена |
|||||||
ненаследственное. |
 |
случае |
активность |
гена-супрессора |
|||
наследственной формы множественные |
опухолеобразования.Поэтомугетерозиготы |
||||||
опухоли обычно возникают независимо в |
по мутации данного гена являются |
||||||
обоих глазах. При ненаследственной |
генетически предрасположенными к |
||||||
форме, только одна опухоль возникает |
опухолеобразованию. |
Первая |
æå |
||||
только в одном глазу. |
|
|
|
соматическая мутация этого локуса, делает |
|||
|
|
|
|
данную клетку гомозиготой по мутации, из |
|||
|
|
|
|
|
|
|
20-10 |