

Внутреннее строение птиц Общее строение систем и органов
Пищеварительная система птиц имеет ряд особенностей, которые связаны, прежде всего, с полётом и большой потребностью в энергии: роговой клюв, отсутствие зубов, наличие зоба, два отдела желудка, слабая дифференцировка заднего отдела кишечника, высокая скорость переваривания пищи.
Видовая специфика пищеварительной системы определяется типом питания и способом захватывания корма. Большинство птиц плотоядны, особенно хищники, которые, благодаря сильной мускулатуре, острым когтям и клюву, могут брать крупную добычу. Их жертвами становятся многие виды позвоночных – млекопитающих (особенно грызунов), рептилий, земноводных и рыб. Некоторые птицы (ястреб-перепелятник) охотятся на других, более мелких видов птиц. Рацион питания многих птиц составляют беспозвоночные – насекомые, черви, моллюски и др. Немало птиц всеядны: помимо животных кормов они потребляют и растительные (ягоды, семена, орехи).
К числу птиц, поедающих преимущественно растительные корма, относятся гуси, некоторые виды куриных, пастушковых, однако и они при случае питаются мелкими беспозвоночными животными. Ряд птиц получают пищу путём процеживания воды, где содержатся планктонные организмы, водоросли и детритные остатки (фламинго, утки).
При недостатке привычных кормов птицы способны переключаться на другие виды пищи, более обильно представленные в данное время. Так, при массовом выселении саранчи многие птицы (утки, чайки, цапли, кулики и др.) истребляют её в больших количествах, чем приносят большую пользу природным сообществам и человеку, выступая природным регулятором численности быстро размножающихся видов.
Основной орган захвата пищи при любом типе питания является клюв, имеющий разную форму (см. внешнее строение птиц).
В ротовой полости имеется язык с хорошо развитой мускулатурой и часто снабжённый роговыми шипиками для удержания добычи. Хищные птицы, проглатывающие крупные куски добычи, имеют короткий язык. У птиц с фильтровальным ротовым аппаратом язык помогает отделять воду, частицы грунта от пищи. Язык зерноядных птиц способствует удержанию плотных семян при их раскусывании. Дятлы имеют язык с крючковидными выростами и способный сильно выдвигаться при вытаскивании насекомых и их личинок из-под коры. Птицы, питающиеся нектаром цветков (колибри, нектарницы), снабжены тонким шиловидным клювом с сосущей трубочкой внутри. У ряда птиц (пеликаны, вороновые, вьюрки и др.) под языком имеется углубление, позволяющее резервировать корм на некоторое время (например, для её переноса).
Для смачивания пищи служит слюна, поступающая по протокам из слюнных желез. У некоторых птиц (стрижи, ласточки) слюна быстро затвердевает на воздухе и используется птицами для скрепления материалов при гнездостроении. Слюна некоторых птиц содержит фермент амилазу для первичной химической обработки пищи.
За корнем языка находится гортанная щель – входное отверстие в трахею и начало длинного пищевода, имеющего гладкую мускулатуру. У ряда птиц (голуби, хищники, куриные, попугаи) в нижней трети пищевода образуется зоб (рис. 63), где пища временно скапливается и смачивается. Стенки зоба некоторых птиц (попугаи, голуби) вырабатывают «молочко», содержащее около 10% белка и 12–15% жира и используемое для вскармливания птенцов.
|
|
Рис.63. Схема пищеварительной системы голубя: 1 – пищевод, 2 – зоб, 3 – железистый желудок, 4 – мускульный желудок, 5 – двенадцатиперстная кишка, 6 – поджелудочная железа, 7 – печень, 8 – желчные протоки, 9 – селезенка, 10 – петли тонкой кишки, 11 – прямая кишка, 12 – клоака, 13 – парная слепая кишка. ЗАРИСОВАТЬ |

Желудок птиц состоит из двух отделов – железистого и мускульного, наделённых разными функциями. В железистом желудке, который выглядит как расширение конца пищевода, пища смачивается и подвергается ферментной обработке слизистого секрета, выделяемого железами стенок.
Следующий за ним мускульный желудок имеет толстые стенки, изнутри покрытые роговидной кутикулой, предохраняющей от механических повреждений при перетирании пищи. Процесс механического размельчения пищи осуществляется, благодаря сокращениям мощной мускулатуры стенок.
Для более успешного перетирания грубой пищи (семена, зёрна) ряд птиц заглатывает мелкие камешки (гастролиты), которые служат своеобразными жерновами. При этом развивается давление, равное примерно 20–30 кг на кв. см. Наличие такого способа размельчения пищи у птиц компенсирует отсутствие зубов.
Хищные птицы, поглощающие животных с шерстью, хитиновыми образованиями, отрыгивают подобные неперевариваемые остатки из желудка в виде «погадок», чтобы не засорялся кишечник. Форма таких погадок носит видоспецифический характер – по ним можно установить вид кормившейся птицы.
За желудком следует тонкий кишечник, началом которого является изгиб двенадцатиперстной кишки, в которую впадают протоки поджелудочной железы, закреплённой на брыжейке. Сюда же открываются и желчные протоки, отходящие от желчного пузыря двухлопастной печени (у некоторых птиц желчный пузырь отсутствует, например, у голубя). Тонкий кишечник образует несколько петель и переходит в короткую прямую кишку, имеющую анальное отверстие в клоаку. (У птиц отсутствует подразделение на толстый и прямой отделы кишечника).
На границе тонкой и прямой кишки имеют место небольшие слепые выросты, предназначенные для процессов брожения с помощью микрофлоры, поэтому они более выражены у растительноядных птиц. В целом, кишечник птиц гораздо длиннее, чем у рептилий: он превышает длину тела примерно в пять–десять раз и более. Высока и интенсивность переваривания пищи: мягкие животные корма (гусеница) перевариваются за 10–15 минут. Твёрдая пища (зёрна) требует более продолжительной обработки – порядка 3–4 часов.
На спинной стороне клоаки птенцов формируется фабрициева сумка, выполняющая функции кроветворения (белые кровяные клетки) и клеточного иммунитета (у взрослых птиц она подвергается редукции).
Дыхательная система и механизмы дыхания птиц отличаются рядом особенностей, которые, прежде всего, определяются полётом, требующим значительного количества энергии и кислорода для окислительных процессов. Специфика органов дыхания заключается в значительном усложнении строения дыхательных путей и разнице в механизмах дыхания при разной двигательной активности птиц.
Воздух через наружные ноздри, носовые ходы и хоаны попадает в ротовую полость. Через гортанную щель проходит в верхнюю гортань, которая поддерживается системой хрящей (непарным перстневидным и парными черпаловидными). Следующая за гортанью трахея представляет собой гибкую трубку, стенки которой содержат хрящевые кольца. В месте ответвления от трахеи двух бронхов находится нижняя гортань с голосовым аппаратом, определяющим звукоиздавание. Он состоит из голосовых перепонок, натянутых между последними кольцами трахеи и полукольцами бронхов, и голосовых мышц, сокращение которых вызывает изменение положения перепонок при прохождении воздуха.
У некоторых птиц издаваемые звуки усиливаются за счёт особого образования, представляющего собой тонкостенный барабан, формирующийся при разрастании нижней части трахеи. Изменение силы звуков у ряда птиц (журавли) может осуществляться с помощью петель трахеи.
Лёгкие птиц небольшие по размеру, заполнены губчатой тканью и задней частью прикреплены к рёбрам вблизи позвоночника. При входе в лёгкие бронхи распадаются на 15–20 вторичных бронхов, которые соединяются между собою парабронхами, дающими ещё более мелкие ветвления – бронхиоли. Каждое из этих образований представляет собой ячеистый вырост, густо оплетённый капиллярами кровеносной системы, куда поступает кислород.
Большинство вторичных бронхов оканчиваются слепо в лёгких. Часть из них выходят за пределы лёгочной ткани, образуя воздушные тонкостенные мешки. Воздушные мешки расположены между внутренними органами: две–четыре пары шейных, непарный межключичный, две–три пары грудных и пара крупных брюшных.
Межключичный и брюшные мешки дают ответвления в полые кости плеча и бедра соответственно. Они играют роль в облегчении массы тела, предохраняют от перегрева его полость, служат своеобразными амортизаторами для защиты внутренних органов от механических повреждений во время взлёта и посадки. Однако основное их значение – в определении механизма дыхания при активном передвижении, особенно в полёте.
Сидящая птица дышит за счёт ритмичного сокращения межрёберной мускулатуры и движения грудной клетки. Опускание грудины вниз приводит к расширению объёма грудной полости, и воздух проходит в лёгкие. Обратное её движение приводит к выдоху. При этом роль насосов выполняют и лёгочные мешки, которые то расширяются, то сужаются, благодаря связи с межрёберной мускулатурой.
При движении по земле, лазании по ветвям дополнительным насосом служат воздушные мешки, приводимые в действие мускулатурой задних конечностей. У летящей птицы грудная клетка, благодаря напряжениям мышц крыла, фиксируется неподвижно, и роль механизма нагнетания воздуха выполняют воздушные мешки. Они приводятся в движение мускулатурой крыльев: при их взмахе воздушные мешки растягиваются, и воздух быстро входит в лёгкие. При опускании крыльев производится выдох. Объём воздушных мешков значительно превышает ёмкость лёгких, однако газообмен в них не идёт.
Для птиц характерно такое явление как «двойное дыхание». Оно заключается в том, что окисление крови в лёгких осуществляется дважды за один акт дыхания – на вдохе и выдохе. Происходит это следующим образом. При вдохе воздух из лёгких насасывается в передние воздушные мешки, а воздух, поступивший из внешней среды по трахее, бронхам быстро проходит в лёгкие и задние воздушные мешки (задние грудные и брюшные). При этом в лёгких осуществляется газообмен.
При выдохе воздух, содержащий ещё значительное количество кислорода, выталкивается из этих мешков в лёгкие, где снова идёт газообмен. Одновременно воздух из передних мешков (шейных, межключичного и переднего грудного), содержащий углекислоту, выдавливается наружу.
Более полному насыщению крови кислородом способствует также движение крови по лёгочным сосудам навстречу току воздуха («принцип противотока»). Частота дыхания зависит от скорости полёта: чем больше взмахов крыльев, тем она больше и, соответственно, выше содержание поступающего в кровь кислорода, обеспечивающего процессы окисления и выхода энергии.
Кровеносная система и кровообращение птиц, по сравнению с рептилиями, имеют значительные отличия: артериальная и венозная кровь не смешиваются, благодаря полному разделению малого и большого кругов кровообращения. Сохраняется лишь одна (правая) дуга аорты. В почках частичная редукция воротной системы.
Особенности в системе кровоснабжения во многом определяются строением сердца, которое полностью разделено на две части – левую с артериальной кровью и правую с венозной. Сверху сердце покрыто околосердечной сумкой и состоит из четырёх камер – двух тонкостенных предсердий и двух желудочков, имеющих толстые стенки и сильную поперечнополосатую мускулатуру.
Малый круг кровообращения начинается с правого желудочка, из которого легочная артерия, подразделяясь на правую и левую части, несёт венозную кровь для окисления в лёгкие. Окисленная кровь по лёгочным венам изливается в левое предсердие, а затем проходит в левый желудочек.
Большой круг кровообращения берёт начало с левого желудочка, от которого отходит единственная, правая дуга аорты, несущая артериальную кровь. Отделив крупные безымянные артерии (левую и правую), дуга аорты в виде спинной аорты тянется вдоль позвоночника (рис. 65).
От каждой безымянной отходит по паре общих сосудов – сонная, идущая к голове и подключичная, которая, в свою очередь, отсоединяет плечевую и грудную артерии, снабжающие кровью мышцы крыла и грудной клетки. От спинной аорты отходят сосуды, доставляющие артериальную кровь к внутренним органам, задним конечностям, мышцам брюшной стенки и органам тазовой области. К их числу относятся: непарные – внутренностная, брыжеечная, хвостовая и парные – почечные, бедренные, подвздошные и седалищные артерии.
Венозная система во многом схожа с таковой у рептилий. Из головы венозная кровь собирается в парные яремные вены (рис. 66). Объединяясь с плечевой и грудной венами, они образуют крупные передние полые вены, по которым венозная кровь изливается в правое предсердие. Венозная кровь от задней части тела отходит по хвостовой и парным внутренним подвздошным венам. Сливаясь, они дают начало трём крупным венам – непарной копчиково-брыжеечной и парным воротным венам почек.
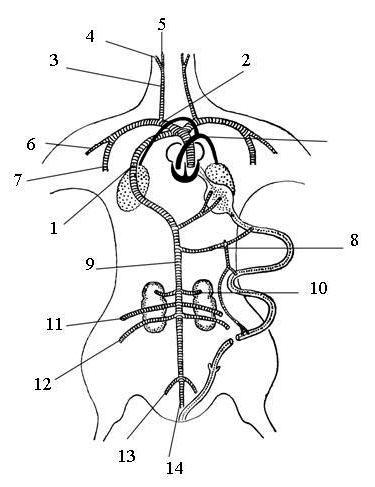
Рис. 65. Схема артериальной системы птицы:
1– правая дуга аорты, 2 – правая безымянная артерия, 3 – правая общая сонная артерия, 4 – наружная сонная артерия, 5 – внутренняя сонная артерия, 6 – подключичная артерия, 7 – правая грудная артерия, 8 – кишечная артерия, 9 – спинная аорта, 10 – левая почечная артерия, 11 – правая бедренная артерия, 12 – правая седалищная артерия, 13 – правая подвздошная артерия, 14 – хвостовая артерия, 15 – легочная артерия (черным цветом обозначены артерии с венозной кровью).
Воротная система почек, по сравнению с таковой у рептилий, частично редуцирована. Это проявляется в том, что только часть крови расходится по почечным капиллярам почек; остальная кровь идёт по общим подвздошным венам, вбирающим кровь из парных бедренных и седалищных вен. Общие подвздошные вены, сливаясь с воротными почечными венами, по выходе из почек формирует заднюю полую вену, несущую кровь в правое предсердие. Копчиково-брыжеечная вена, вбирая кровь из нескольких сосудов, идущих от желудка и кишечника, образует воротную систему печени. Воротные вены печени впадают в заднюю полую вену.
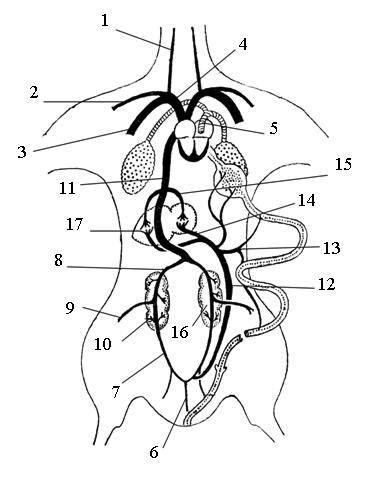
Рис. 66. Схема венозной системы птицы:
1 – правая яремная вена, 2 – правая подключичная (плечевая) вена, 3 – правая грудная вена, 4 – правая передняя полая вена, 5 – легочная вена, 6 – хвостовая вена, 7 – правая воротная вена почек, 8 – правая почечная вена, 9 – правая бедренная вена, 10 – правая подвздошная вена, 11 – задняя полая вена, 12 – копчиково-брыжеечная вена, 13 – надкишечная вена, 14 – воротная вена печени, 15 – левая почечная вена, 16 – левая почка, 17 – печень (заштрихованы вены с артериальной кровью)
Скорость циркуляции крови определяется работой крупного сердца и частотой его сокращений. Объём крови, содержание в ней эритроцитов, гемоглобина и кислородная ёмкость значительно выше (в 2–4 раза), чем у пресмыкающихся. Это отражает высокий уровень обменных процессов, обеспечивающих необходимое количество энергии.
Выделительная система взрослых птиц представлена, как у всех амниот, тазовыми почками (метанефрос).
На эмбриональной стадии закладываются мезонефричекие почки, которые впоследствии редуцируются. Тазовые почки птиц отличаются довольно крупными размерами и в виде трёх долей (рис. 67) залегают в углублениях сложного крестца. Над ними располагаются надпочечники – железы внутренней секреции. От почек отходят мочеточники, впадающие в клоаку. Мочевой пузырь у птиц отсутствует. Основным продуктом белкового распада является мочевая кислота, которая выделяется в виде кашицеобразной массы.
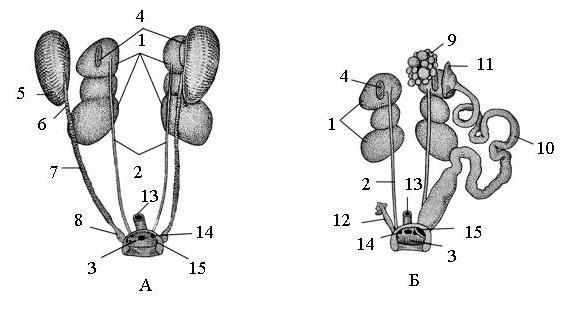
Рис. 67. Мочеполовая система голубя. А – самец; Б – самка:
1 – почка, 2 – мочеточник, 3 – полость клоаки, 4 – надпочечник, 5 – семенник, 6 – придаток семенника, 7 – семяпровод, 8 – семенной пузырек, 9 – левый яичник, 10 – левый яйцевод, 11 – воронка яйцевода, 12 – остаток редуцированного правого яйцевода, 13 – прямая кишка, 14 – моче-выделительное отверстие, 15 – половое отверстие.
Образование мочи в почках происходит в основном за счёт работы железистых клеток, находящихся в стенках почечных канальцев, оплетённых капиллярами воротной системы. Наряду с этим, идёт фильтрование крови через мальпигиевы клубочки боуменовых капсул. Образовавшийся первичный фильтрат подвергается обратному всасыванию в нижней части нефридиальных трубочек, где образуются их изгибы – петли Генле. В результате процесса реабсорбции моча выделяется в концентрированном виде, организм сохраняет воду и полезные для него вещества.
У многих видов птиц, особенно морских и ряда пустынных, имеются носовые железы, способствующие выделению избытка солей из организма. Они располагаются в орбите глаз и имеют протоки в носовые ходы. Концентрация солей в секрете желёз в 4–5 раз выше, чем в крови. Наличие солевых желёз позволяет экономить воду при её дефиците в месте обитания и даже пить солёную воду.
Половая система и размножение птиц имеют большое своеобразие.
Птицы размножаются только кладкой яиц, которые насиживают в течение определённого времени, а по выходе птенцов выкармливают их, проявляя сложные поведенческие реакции, связанные с заботой о потомстве.
Семенники самцов в виде парных овальных тел расположены вблизи верхней доли почек и подвешены на брыжейке (рис. 67 А). Их объём резко возрастает ко времени размножения. С внутренней стороны семенников находятся придатки семенника (остатки мезонефроса), от которых отходят парные семяпроводы, в нижней части имеющие семенные пузырьки для резервирования сперматозоидов. Семяпроводы гомологичны вольфовым каналам и открываются половыми отверстиями в клоаку. Совокупительные органы в виде непарного выроста клоаки имеются лишь у ряда систематических групп птиц (тинаму, страусы, гусеобразные).
Половая система самки представлена только левым яичником и соответствующим яйцеводом (рис. 67 Б). Правые яичник и яйцевод у подавляющего большинства птиц редуцированы (при наличии и правого яичника функционирует только один яйцевод). Яичник неправильной формы, имеет зернистую структуру и также подвешен на брыжейке вблизи переднего края левой почки. Яйцеводом служит мюллеров канал, который расширенной воронкой открывается около яичника.
Созревшая яйцеклетка выходит через разрыв фолликула в полость тела и подхватывается воронкой яйцевода. Оплодотворение яйцеклетки происходит в верхней части яйцевода. Проходя по яйцеводу, зигота последовательно покрывается защитными оболочками, которые формируются за счёт выделений особых желёз стенок яйцевода. Формирование яйца занимает разный промежуток времени (12–48 ч.) в зависимости от видовых особенностей птицы.
Развивающийся эмбрион находится на желтковой части яйца (зародышевый диск) и окружён целым рядом оболочек. Желток вместе с зародышевым диском ограничивает желтковая оболочка, которая была сформирована при прохождении зиготы по начальному отделу яйцевода. За ней следуют белковая оболочка и две тонких пергаментных оболочки (подскорлуповые), которые на тупом конце яйца расходятся с образованием воздушной камеры (запас воздуха). Между желтком и внутренней подскорлуповой оболочкой крепится халаза – жгутообразное белковое образование, благодаря которому при любом движении яйца зародышевый диск всегда оказывается наверху.
Развивающийся зародыш, как у всех амниот, окружён особыми зародышевыми оболочками (амниотическая, серозная, аллантоис), которые обеспечивают не только дополнительную защиту, но и процессы его жизнедеятельности – питание, дыхание и выделение. Главным источником энергии, необходимой для нормального развития эмбриона, является желток. Белковая оболочка служит дополнительным энергетическим резервом и источником воды.
Соотношение желтка и белка в яйцах разных видов значительно варьирует. У воробьиных птиц желток может составлять 10–25%, а белок – 70–80% от массы яйца. Скорлупа формируется в основном из углекислого кальция (92–95%) с добавлением других химических веществ (углекислого магния, фосфорнокислого кальция, фосфорнокислого магния и небольшого количества органических веществ).
Продолжительность инкубационного периода во многом определяется размерами птицы и яиц, местом размножения, типом гнезда, интенсивностью насиживания. Самый короткий период выведения птенцов имеет место у мелких воробьиных птиц (11–14 суток). Птицы среднего и крупного размеров насиживают яйца более продолжительное время: 17–20 суток (вороновые); 20–40 суток (гусеобразные); около двух месяцев (альбатросы, грифы, крупные пингвины).