

Розділ 8. Основи молекулярної генетики
8.1. Біосинтез нуклеїнових кислот
8.1.1. Реплікація ДНК. Найбільш важливі функції нуклеїнових кислот - це участь у передачі спадкових ознак, у процесі біосинтезу білка та його регуляції. Основним носієм генетичної інформації у більшості організмів є ДНК. Винятком є тільки окремі фаги і віруси, в яких носієм спадкової інформації служать молекули РНК. Комплементарність ланцюгів ДНК становить хімічну основу важливої функції ДНК - збереження і передача спадкових (генетичних) ознак.
Модель подвійної спіралі ДНК, запропонованої в 1953 році Дж. Уотсоном та Ф. Кріком, не тільки описує структуру ДНК, але й показує механізм подвоєння – реплікацію ДНК – рівний розподіл між дочірніми клітинами генетичного матеріалу, що має важливе біологічне значення у процесі збереження і передачі генетичної інформації.
З
Рис.8.1. Біосинтез ДНК з
врахуванням комплементарності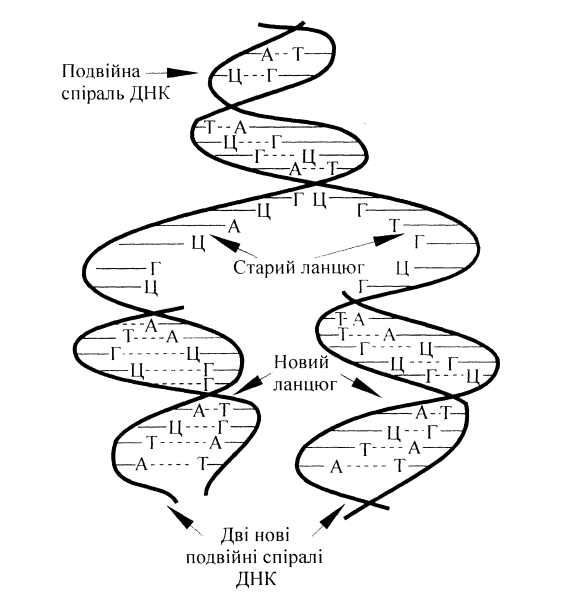
При поділі клітини подвійна спіраль ДНК розкручується і розподіляється на два ланцюги. На кожному ланцюзі, як на матриці, проходить біосинтез нового ланцюга ДНК з врахуванням принципу комплементарності(рис.8.1).Утворений полінуклеотидний ланцюг не ідентичний, але комплементарний вихідній матриці.
У результаті утворюються дві нові подвійні спіралі, кожна з яких включає один старий (материнський) і один новий - синтезований (дочірний) ланцюг. Такий процес точного копіювання молекули ДНК, в результаті якого утворюються дві однакові двоспіральні молекули, називається реплікацією. Таким чином, спадкові ознакизберігаються в поколіннях. Реалізація генетичної інформації здійснюється за рахунок передачі закодованої в ДНК інформації молекулам мРНК (транскрипції) та подальшого розшифрування цієї інформації при синтезі білка (трансляції).
Аналогічно в ядрі відбувається синтез молекули мРНК, яка потім служить матрицею для біосинтезу білка в цитоплазмі. Утворена мРНК комплементарна цій ділянці ДНК, на якій вона синтезувалася. При цьому аденіновій основі в ДНК буде відповідати урацилова основа в РНК, а пентозним залишком у ланцюзі РНК буде виступати рибоза. Синтез мРНК по суті є переписуванням (транскрипцією) генетичної інформації з ділянок (генів) ДНК на мРНК. Остання потім використовується як матриця, на якій в присутності рибосом і відповідних тРНК, що переносять залишки -амінокислот, проходить біосинтез білків.
Генетична інформація записана (закодована) в нуклеотидній послідовності ДНК. Три нуклеотиди, які контролюють включення певної амінокислоти в білок у процесі його біосинтезу, називають кодоном.
Виключно велике значення нуклеотидів не обмежується тільки цим, що вони є у складі нуклеїнових кислот. Окремі нуклеотиди входять у склад небілкової частини (кофермента) ферментних систем і приймають участь в обміні речовин. Важливу групу коферментів становлять не тільки монофосфати (АМФ, ГМФ, ЦМФ тощо), але й нуклеозидполіфосфати (АДФ, АТФ). НАД+, НАДФ+, ФАД, ФМН беруть участь у реакціях біологічного окиснення; УТФ, УДФ – у реакціях біосинтезу глікогену; ЦТФ, ЦДФ – у біосинтезі гліцерофосфоліпідів. У процесі обміну речовин в організмі, як універсальне джерело і акумулятор енергії, використовуються АТФ, ГТФ тощо.
Для синтезу полімерної молекули ДНК потрібна енергія. Джерелом енергії в реакціях полімеризації мононуклеотидів є енергія, що звільняється дезоксирибонуклеозидтрифосфатами, які беруть участь у синтезі ДНК. Пірофосфат, що утворюється під дією пірофосфатази, також розщеплюється на дві молекули ортофосфату з утворенням додаткової енергії для біосинтезу ДНК.
Біосинтез ДНК вимагає наявності специфічних ферментів, що каталізують окремі етапи синтезу, і безлічі білкових чинників, необхідних для регулювання процесу реплікації та прояву каталітичної активності ферментів.
У реплікації ДНК, що включає розпізнавання точки початку процесу, розплітання материнських ланцюгів ДНК у реплікативній вилці, ініціацію біосинтезу дочірніх ланцюгів та подальшу їх елонгацію і, нарешті, закінчення (термінація) процесу, бере участь більше, ніж 40 ферментів й білкових чинників, об’єднаних в єдину ДНК-репліказну систему.
(дНМФ)n + дНТФ → (дНМФ) n+1 + ФФн
ДНК ДНК подовжена
ДНК – полімераза, що здійснює утворення 3’-5’ фосфодіефірних зв’язків, могла функціонувати тільки в присутності вже існуючої („передіснуючої”) ДНК. Ця ДНК виконує функції:
- затравки для подовження ланцюга полідезоксирибонуклеотиду;
- матриці, що визначає послідовність включення в ланцюг нових нуклеотидів.
ДНК – полімераза І не може синтезувати повністю новий ланцюг, а спроможна приєднувати дНМФ до вже існуючого ланцюга. Синтез нового ланцюга ДНК відбувається в напрямку 5’- 3’, тобто ДНК – полімераза послідовно приєднує нуклеотиди (дНМФ з наявних дНТФ) до 3’ – кінця одного з ланцюгів ДНК (ланцюг „затравки”). Для синтезу нового ланцюга ДНК необхідний ланцюг – матриця, нуклеотиди сполучаються з 3’- кінцем ланцюга „затравки” відповідно до нуклеотидної послідовності в ланцюгу „матриці” (за принципом комплементарності).
Механізм подовження (елонгації) ланцюга в ДНК полягає в утворенні нових 3’-5’ фосфодіефірних зв’язків, що синтезуються в напрямку 5’- 3’.
Експериментальне доведення генетичної ролі ДНК (феномен трансформації). Англійський мікробіолог Ф. Гріффіт вивчаючи два штами пневмококівStreptococcuspneumoniae, що здатний викликати пневмонію в людини та мишей (капсульноїS-форми) та непатогенного мутанта (безкапсульноїR-форми), відкрив явищетрансформації, що полягало в можливості перетворення непатогенноїR-форми в патогеннуS-форму:
трансформація
R → S
Гріффіт Ф. встановив, що трансформація пневмококів відбувається за умов взаємодії в організмі піддослідних тварин вбитої нагріванням S-форми (патогенної) з живою непатогенноюR-формою. У вбитих нагріванням вірулентних клітинах пневмококів (штамS) присутній певнийтрансформуючий фактор , який проникаючи в живі невірулентні клітини (штамR) змінює біологічні властивості останніх, надає їм властивість патогенності, ця властивість є спадковою:
трансформуючий фактор
R → S
Пізніше групою вчених США було доведено, що цим трасформуючим фактором пневмококів є молекули ДНК.
ДНК
R → S
Отже, поняття "фактор спадковості", або "ген"набуло конкретного молекулярного змісту – це молекула ДНК.
8.1.2. Напівкосервативний та консервативний механізми реплікації ДНК. М. Мезельсон і Ф. Сталь (1957 р) показали напівконсервативний механізм реплікації ДНК, що включає утворення дочірніх молекул ДНК, в кожній з яких зберігається лише один материнський ланцюг (рис. 8.2).
Д
Рис. 8.2. Напівконсервативний
механізм реплікації ДНК
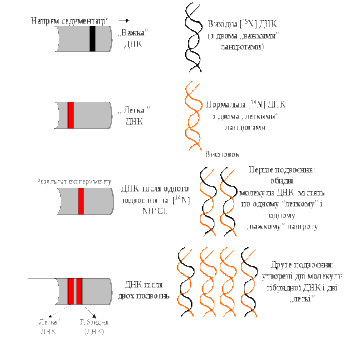
Експеримент включав наступні етапи:
- вирощування клітин E. Coli на середовищі, що містило як джерело азоту NH4CI з легким ізотопом (14N); була встановлена щільність „легких” молекул ДНК, які містилися в таких клітинах;
- вирощування клітин E. сoli протягом декількох поколінь на середовищі, що містило як джерело азоту NH4CI з важким ізотопом (15N); була встановлена щільність „важких” молекул ДНК, які містилися в таких клітинах;
-
Рис. 8.3. Механізми реплікації
ДНК: А – консервативний, Б –
напівконсервативний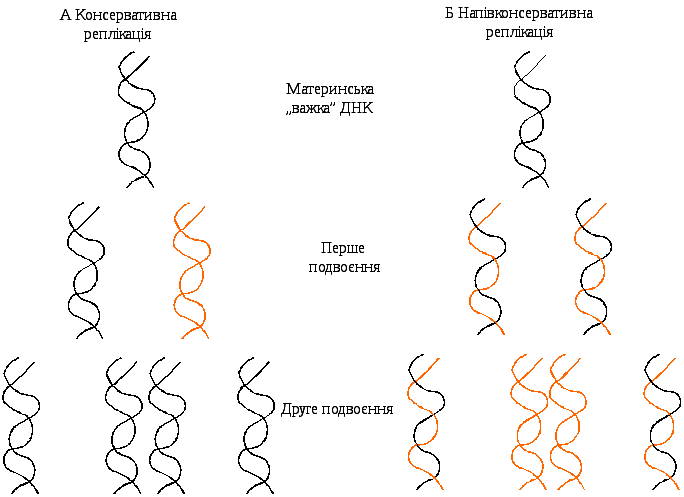
Крім зазначеної моделі Уотсона і Кріка, теоретично можливий і інший механізм – консервативний, при якому заново синтезований нуклеотидний ланцюг утворюється на подвійній спіралі ДНК (рис.8.3). Материнська ДНК залишається незайманою, а з вільних мононуклеотидів складаються повністю нові молекули ДНК.
8.1.3. Послідовність етапів і ферменти реплікації ДНК у прокаріотів. Процес реплікації ДНК у прокаріот складається із трьох основних етапів: ініціація (початок), елонгація (подовження ланцюга) і термінація (кінець утворення дочірніх ДНК). Кожна стадія відбувається за участю відповідних ферментів і білкових факторів.
1. Розплітаючі білки розривають водневі зв’язки між комплементарними основами подвійної спіралі ДНКзб утворенням “реплікативної вилки”. Характерним є те, що подальше переміщення реплікативної вилки можливе тільки при розкручуванні материнських ДНК і одночасному синтезі обох нових ланцюгів ДНК,
2. Затравочна ДНК-залежна РНК-полімераза (РНК-полімераза або праймаза), що
бере участь у стадії ініціації реплікації ДНК та каталізує синтез короткого олігорибонуклеотиду (від 10 до 60 нуклеотидів), тобто праймера, з якого потім починається синтез ДНК. Праймази розрізняються як за структурою, так і за специфічністю дії. /
Праймасома представлена ансамблем із 7 різних субодиниць, що включають близько 20 поліпептидів загальною молекулярною масою 70 000 Да. За допомогою спеціального білка праймасома піддається швидкому переміщенню до відстаючого ланцюга ДНК за рахунок енергії, що генерується АТФазною активністю спеціального білка. До складу праймасоми входить також комплекс білків dnа B і dna C, який поблизу вилки реплікації періодично бере участь у формуванні специфічної вторинної структури ДНК, відповідної для розпізнавання праймазою.
3. ДНК–полімерази. В прокаріот у процесі реплікації беруть участь три форми ДНК- полімераз (І, ІІ, ІІІ). Усі вони проявляють два види активності: полімеразну- зшивають дезоксирибонуклеотиди3’,5’-фосфодіефірними зв’язками і нуклеазну - гідролізують фосфодіефірні зв’язки.
а) ДНК – полімераза І виконує такі функції:
- 5’ – 3’ – полімеразна активність;
- 5’ – 3’- екзонуклеазна активність (здатність видаляти нуклеотиди вище від напрямку синтезу);
3’ – 5’ екзонуклеазна активність („коригуюча” активність, або спроможність видаляти вже включений нуклеотид, якщо він включений помилково;
ДНК-полімераза І каталізує відщеплення затравочного олігорибонуклеотидного праймера і заміну його дезоксирибонуклеотидами.
б) ДНК полімераза ІІ, основна функція – участь у репарації ДНК;
в) ДНК – полімераза ІІІ головний – фермент, що реалізує процес елонгації ДНК у E.соli.
Основним ферментом, що каталізує біосинтез новоутвореної ДНК (точніше, стадію елонгації реплікації ДНК), є ДНК-полімераза III.
ДНК-полімераза III в Е. соli складається з 10 субодиниць. У димерній формі ДНК-полімераза III каталізує синтез лідируючого і відстаючого ланцюгів ДНК при реплікації. Фермент має 5’ – 3’ – полімеразну, 3’ – 5’ – екзонуклеазну та АТФазну активності.
ДНК – полімераза ІІІ може каталізувати сполучення в ланцюг багатьох тисяч мононуклеотидів (на відміну від 15 – 20 нуклеотидів при дії ДНК – полімерази І).
4. Рибонуклеаза Н. Бере участь у гідролізі РНК-затравки разом із ДНК полімеразою І.
5. ДНК-лігази – зшиваючі ферменти. Вони беруть участь у процесі сполучення одне з одним новосинтезованих фрагментів ДНК з утворенням фосфодіефірних зв’язків.