

Lektsii-biologia
.pdfДанные о видовой специфичности строения ДНК получены в начале 50-х годов Эрвином Чаргаффом и его сотрудниками. Лю - бая ДНК состоит из четырех типов нуклеотидов, содержащих одно из четырех азотистых оснований: аденин, тимин, гуанин, цитозин. Чаргафф с сотрудниками обнаружил, что соотношения между нуклеотидами, входящими в состав ДНК одинаковы в пре - делах вида и различаются у разных видов. Кроме того, число аде -
ниловых нуклеотидов равно числу тимидиловых, а число гуани |
- |
ловых равно числу цитозиловых. Это открытие послужило глав |
- |
ным ключом к выявлению структуры молекулы ДНК. |
|
Трансформация у бактерий – это передача от одной бактери |
- |
альной клетки другой наследственного материала. В 1928 году |
|
Фред Гриффит изучал трансформацию у двух штаммов пневмо |
- |
кокка: вирулентного (вызывающего гибель мышей) и авирулент |
- |
ного (не вызывающего гибели). |
|
Если убить вирулентные бактерии и вводить их вместе с жи |
- |
выми авирулентными, то часть животных все равно погибнет. |
|
Этот опыт свидетельствовал о том, что часть генетического мате -
риала из вирулентных бактерий проникла в авирулентные и сде |
- |
лала их вирулентными. После того как вирулентные клетки раз |
- |
делили на отдельные компоненты: углеводы, липиды, белки и |
|
ДНК, было обнаружено, что именно ДНК наделило живые бакте - рии генетическим признаком, которого у них до этого не было.
Трансдукция – это явление, заключающееся в том, что вирусы, выходя из бактериальной клетки, в которой они паразитировали, могут захватывать с собой часть ДНК и, перемещаясь в новые клетки передавать им признаки прежних хозяев.
Так, при внесении в неподвижные бактерии вирусов, размножав -
шихся в клетках подвижных культур, некоторые из бактерий ста |
- |
новятся подвижными. |
|
Половой процесс у бактерий – коньюгация, когда происходит |
|
обмен молекулами ДНК, и хотя новые бактерии не образуются, |
|
но их наследственный материал изменяется, т.к. происходит ре |
- |
комбинация генетического материала. |
|
Строение вируса или бактериофага : вирус состоит из молеку -
лы ДНК (или РНК), заключенной в белковую оболочку. С помо - щью радиоизотопов было доказано, что именно ДНК проникает в
91D
клетку, а белковая оболочка остается снаружи. В результате обра - зуются новые фаговые частицы. Таким образом, ДНК является носителем генетической информации и дает начало новым бакте - риофагам.
Косвенные доказательства : в соматических клетках разных органов содержится одинаковое количество ДНК, вдвое большее, чем в половых. Количество белков варьирует и не всегда больше, чем в половых.
2).Следующий этап развития молекулярной генетики связан с таким важным открытием, как установление структурной органи - зации молекулы ДНК– в 1953 году. Крик и Уотсон установили, что ДНК состоит из двух спирально-закрученных цепей.
В настоящее время принято говорить о первичной, вторичной и третичной структуре ДНК: Первичной структурой ДНК называют линейную полинуклеотидную цепь, в которой мононуклеотиды соединены 3`,5`- фосфоэфирными связями. Модель Крика и Уот - сона – это вторичная структура ДНК, основанная на принципе комплементарности и получившая название двойной спирали. Один виток спирали состоит из 10 нуклеотидов, размер витка 3,4 нм.
Дополнительное структурирование в пространстве двуспираль - ной молекулы ДНК (в конденсированных хромосомах) с образо - ванием суперспирали является третичной структурой.
3).Определение направления передачи информации – «один ген
–один фермент» - Бидл и Татум – 50-е годы. (В настоящее время
–один ген – один полипептид).
4).Расшифровка генетического кода – Ниренберг, Очоа (К 1964 году расшифрованы коды для всех аминокислот). Выяснение ме - ханизма экспрессии генов у прокариот и ее регуляции – Франсуа Жакоб и Жан Моно – 50-е годы.
5).70-е годы и до настоящего времени – выявление особенно - стей экспрессии генов у эукариот. Развитие генетической инже - нерии.
Генетический код и его свойства
92D
Генетический код – это система записи информации о последо - вательности расположения аминокислот в белках с помощью по - следовательности расположения нуклеотидов в молекуле ДНК.
Свойства генетического кода:
1.Код триплетен – каждая аминокислота зашифрована последова - тельностью трех нуклеотидов, называемых триплетом или кодо - ном.
2.Код вырожден – каждая аминокислота кодируется более чем одним кодоном. (Исключения – аминокислоты метионин и трип -
тофан. Они кодируются одним кодоном.) |
|
3.Код универсален – одни и те же триплеты кодируют одни и те |
|
же аминокислоты у всех организмов. |
|
4.Генетический код не перекрывающийся – триплеты нуклеоти |
- |
дов не перекрывают друг друга. У некоторых вирусов обнаруже |
- |
но перекрывание, но это исключение. |
|
5.Имеются стартовый и терминальный кодоны. Стартовый – еди - ный для всех – АУГ. Терминальный трех видов – УАГ, УАА, УГА.
Функционально-генетическая классификация генов
В настоящее время ген рассматривается как единица функцио - нирования наследственного материала. Ген – это участок молеку - лы ДНК, ответственный за синтез одного полипептида.
Различают три вида генов:
-структурные;
-модуляторы;
-регуляторы.
Структурные гены несут информацию об аминокислотах в бел - ках, ферментах, а также о последовательности нуклеотидов в мо - лекулах р-РНК и т-РНК.
Гены-модуляторы влияют на функционирование структурных генов, могут смещать в ту или иную сторону процесс развития признака. Они подразделяются на:
1)ингибиторы или супрессоры (эпистатичные гены),
2)интенсификаторы – могут повышать способность структурных генов к мутациям,
93D
3)модификаторы – могут влиять на структурные гены по типу комлементарности.
Гены-регуляторы контролируют синтез регуляторных белков, а также время включения различных структурных генов в процессе индивидуального развития.
Свойства генов:
1. Дискретность действия – каждый ген действует как самостоя - тельная единица наследственности.
2.Стабильность – при отсутствии мутаций ген передается в ряду поколений в неизмененном виде.
3.Специфичность действия – каждый ген влияет на развитие своего определенного признака.
4.Плейотропия – способность одного гена обеспечивать развитие одновременно нескольких признаков (синдром Марфана).
5. |
Присутствие в виде двух у диплоидных и большего числа ал |
- |
лелей у полиплоидных организмов. |
|
|
6. |
Действие гена дозировано, при изменении числа доз гена в ор |
- |
ганизме изменяется признак (например, при болезни Дауна про |
- |
|
исходит увеличение до трех доз генов 21 хромосомы) |
|
|
Функциональная активность генов или экспрессия ге -
нов
У прокариот она осуществляется в два этапа: транскрипция и трансляция.У эукариот есть еще стадия процессинга.
Экспрессия генов заключается в синтезе на молекуле ДНК мо - лекулы и-РНК, комлементарной ей (или транскрипции – перепи -
сывание, считывание биологической информации) и дальнейшее |
|
ее использование для синтеза белка. Единицей транскрипции в |
|
ДНК является транскриптон, превышающий по размерам струк |
- |
турные гены. Транскриптон в клетках эукариот состоит из неин |
- |
формативной (акцепторной) и информативной зоны. Неинформа - тивная зона начинается геном-промотором (участок из 80 нуклео - тидов), к которому присоединяется фермент РНК-полимераза, ка - тализирующая процесс считывания. У прокариот один вид РНКполимеразы, у эукариот три.
94D
За геном промотором находятся гены-операторы, которые свя -
зывают регуляторные белки (белки, включающие и прекращаю |
- |
щие транскрипцию). |
|
Информативная зона состоит из структурных генов, распола |
- |
гающихся за генами операторами. Структурные гены эукариот |
|
разделены спейсерами – участками ДНК, не несущими информа - ции.
Кроме того, в составе самих структурных генов есть информа - ционные участки – экзоны и неинформационные – интроны. У пркариот спейсеров, экзонов и интронов нет.
В каждой фазе жизненного цикла в клетке транскрибируется только 10% структурных генов, а остальные гены не активны, но часть из них может включаться в других фазах жизненного цикла.
В результате транскрипции у прокариот сразу образуется м- |
|
РНК (зрелая РНК) и сразу же начинается процесс трансляции. |
|
У эукариот транскрибируется большая молекула и-РНК, со |
- |
держащая все неинформативные участки. Она называется РНК- |
|
предшественница или пре-РНК. Поэтому за транскрипцией на |
- |
ступает процессинг, в результате, которого разрушаются все не |
- |
информативные участки: акцепторная зона, спейсеры и интроны, |
|
а оставшиеся экзоны сшиваются (сплайсинг). |
|
На этапе процессинга к начальному отрезку образовавшейся |
|
РНК присоединяется последовательность нуклеотидов, называе |
- |
мая колпачком, к концевому – последовательность остатков аде |
- |
нина – поли- A. |
|
В процессинге происходит модификация нуклеотидов в РНК, |
|
например, их метилирование, гидрирование. |
|
И только после этих превращений образуется зрелая м-РНК, |
|
которая начинается вводной последовательностью, называемой |
|
лидером, и заканчивается концевой последовательностью – трей - лером. Лидер – вводная последовательность нуклеотидов, ком - плементарная последовательности в молекуле р-РНК малой субъ - единице рибосом, которая обеспечивает прикрепление и-РНК к малой субъединице. Трейлер включает нонсенс-кодон и поли-А последовательность.
95D
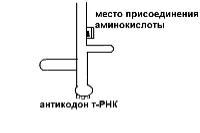
Кодовым элементом м-РНК является триплет нуклеотидов, на - зываемый кодоном. Каждому кодону соответствует определенная аминокислота.
Первичной структурой РНК является, как и в ДНК, линейная цепь полинуклеотидов, в которой мононуклеотиды соединены 3`, 5`-фосфоэфирными связями.
Вторичная структура РНК – изогнутая цепь, а третичная пред - ставляет собой нить, намотанную на катушку, роль катушки игра - ет особый транспортный белок – информатор.
Образующаяся м-РНК идет в цитоплазму к месту синтеза по - липептида (белка), т.е. к рибосомам.
На рибосомах осуществляется процесс трансляции. Трансля - ция – это механизм, с помощью которого последовательность нуклеотидов в молекулах м-РНК переводится в специфическую последовательность аминокислот в полипептидной цепи. Трансляция складывается из трех стадий:
1.Инициация – начало синтеза полипептида.
2.Элонгация – удлинение полипептида.
3.Терминация – окончание синтеза полипептида.
На этапе инициации меньшая субъединица рибосомы узнает стартовый кодон АУГ м-РНК и прикрепляется к ней. АУГ зани - мает первую позицию. После этого присоединяется большая субъединица рибосомы и в ней начинается собственно синтез белка. К большой субъединице подходит т-РНК с аминокислотой.
В молекуле т-РНК одна ее часть присоединяет аминокислоту, а другая – антикодон, спаривается с колоном м-РНК, определяю - щим эту аминокислоту.
96D
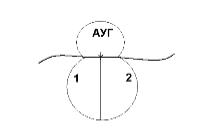
В большой субъединице есть: 1 – аминоацильный участок 2 – пептидильный участок
В т-РНК, антикодон которой комплементарен кодону АУГ м-РНК приносит аминокислоту метианин и останавливается в аминоацильном участке. Стадия инициации контролируется фак - торами инициации. У прокариот их 3, у эукариот 6-7.
Вторая стадия – элонгации начинается с перемещения т-РНК с метианином в пептидильный участок, а на ее место приходит другая т -РНК с новой аминокислотой. Между двумя аминокис - лотами образуется пептидная связь. Тем временем рибосома продвигается вдоль м-РНК, на рибосоме оказывается новый ко - дон, к которому вскоре присоединяется своим антикодоном соот - ветствующая т-РНК.
Все это повторяется многократно, до тех пор, пока рибосома
не дойдет до одного из терминальных кодонов (УАА, УАГ, УГА).
Этим кодонам нет антикодона, нет аминокислоты. Наступает фаза терминации. К одной молекуле м-РНК прикрепляется обыч - но много рибосом и образуются полисомы.
В процессе трансляции наряду с факторами инициации и элон - гации важное значение имеет фермент, связывающий т-РНК с аминоацильным участком – аминоацил т-РНКаза.
Регуляция экспрессии генов у про- и эукариот
97D
У прокариот она осуществляется на уровне промотора, опера - тора
итрансляции.
Уэукариот регуляция происходит на всех этапах экспрессии: транскрипции, процессинга и трансляции.
Подробнее остановимся на регуляции экспрессии генов у бакте - рий.
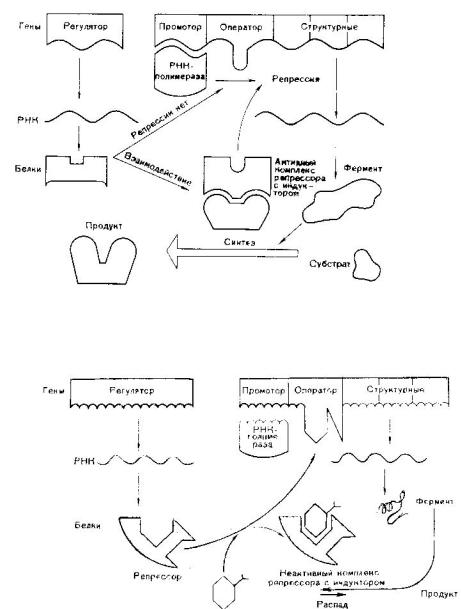
Регуляция экспрессии генов у прокариот
Схема регуляции транскрипции структурных генов прокариоти - ческой клетки по типу репрессии
Схема регуляции транскрипции структурных генов прокариоти - ческой клетки по типу индукции
98D
В 1961 году Жакоб и Моно установили, что у бактерий под контролем гена-регулятора синтезируется белок-репрессор, кото - рый регулирует активность других генов. Белок-репрессор связы - вается с геном-оператором и блокирует его, вследствие чего тран -
скрипция (считывание информации) становится невозможной и |
|
клетка не может синтезировать соответствующие белки. Но если |
|
в клетку проникает какой-либо субстрат, для расщепления кото |
- |
рого необходимо синтезировать ферменты, то происходит следу |
- |
ющее: субстрат присоединяется к репрессору и лишает его спо |
- |
собности блокировать ген-оператор. |
|
К ДНК присоединяются РНК-синтезирующие ферменты и на - чинается транскрипция генов. Синтез белка-фермента прекраща -
ется, если субстрат полностью расщепляется, белок-репрессор |
|
освобождается и снова блокирует гены-операторы. Это пример |
|
регуляции по типу индукции. Также существует регуляция по |
|
типу репрессии: когда конечные продукты биохимической реак |
- |
ции, соединяясь с неактивным белком-репрессором, образуют |
|
комплекс, блокирующий работу гена-оператора. |
|
Антитерминация – заключается в том, что происходит игнори |
- |
рование терминальных кодонов, процесс экспрессии продолжает - ся.
Регуляция у эукариот
1)Регуляция генной активности у эукариот намного сложнее, чем у бактерий. У эукариот она происходит не только на уровне клет - ки. Существуют системы регуляции организма как целого. Огромную роль в регуляции играют гормоны, но регулируют они процессы синтеза белков лишь в клетках-мишенях. Гормоны свя - зываются с белками-рецепторами, расположенными в мембранах таких клеток и включают системы изменения структуры клеточ - ных белков. Те, в свою очередь, могут влиять как на процессы транскрипции, так и процессы трансляции. Каждый гормон через систему посредников активирует свою группу генов. Так адрена - лин включает синтез ферментов, расщепляющих гликоген мышц до глюкозы, а инсулин влияет на образование гликогена из глюко - зы в печени.
99D
2)На стадии транскрипции белки-гистоны участвуют в процессах регуляции генной активности у эукариот. Непременное условие – это деконденсация участка, где происходит транскрипция.
3)Регуляция на уровне трансляции направлена на сохранение ста - бильности м-РНК, а эффективность трансляции осуществляется с помощью факторов инициации, элонгации, терминации.
Лекция 12.
Изменчивость
План
1.Определение и формы изменчивости: модификационная, ком - бинативная, мутационная.
2.Мутагенные факторы.
3.Классификация мутаций.
4.Устойчивость и способы репарации генетического материала.
5.Закон гомологических рядов наследственной изменчивости Н.И.Вавилова.
Определение и формы изменчивости
100D