

Lektsii-biologia
.pdfтому, что передние ноги жирафа оказались длиннее задних, а шея значительно вытянулась. У животных, ведущих подземный образ жизни, орган зрения не использовался и в связи с этим неупраж - нением атрофировался (крот).
Таким образом, Ж.Б.Ламарк считал, что новые признаки всегда полезны и наследуются. Это представление об изначальной целе - сообразности любой реакции на изменение условия, так же как и мнение о прямом воздействии окружающей среды на эволюцион - ные процессы и внутреннем стремлении организмов к прогрессу, оказались ошибочными.
Теория эволюции Ч.Дарвина
В 1858 году Ч.Дарвин и независимо от него А.Р.Уоллес обосновали принцип естественного отбора и представление о борьбе за существование как механизме этого отбора.
Теория эволюции путем естественного отбора основана на сле - дующих положениях. Во-первых, для живого характерно наличие изменчивости, причем для эволюции громадное значение имеет наследственная изменчивость. Вследствие изменчивости призна -
ков и свойств даже в потомстве одной пары родителей почти не |
|
встречается одинаковых особей. При благоприятных условиях |
|
эти различия могут не играть существенной роли, при неблаго |
- |
приятных – каждое мельчайшее различие может стать решающим
в том, останется ли этот организм в живых и даст потомство или |
|
же он будет уничтожен. |
|
Во-вторых, для организмов характерно размножение в геометри |
- |
ческой прогрессии. Потенциально вид в каждом поколении про |
- |
изводит гораздо больше особей, чем их может выжить до взрос |
- |
лого состояния на занимаемой территории. Следовательно, зна |
- |
чительная часть родившихся гибнет в борьбе за существование. В процессе жизнедеятельности каждый организм вступает в много - образные отношения с особями внутри вида, других видов и фак - торами неживой природы. Разнообразные взаимодействия данно - го организма с объектами живой и неживой природы Дарвин и
131D
называл борьбой за существование. Он имел в виду не только жизнь одной особи, но и успех ее в обеспечении себя потомством.
Дарвин выделил три формы борьбы за существовании: внутривидовая (взаимоотношения между особями разного пола, между разными поколениями, отношения в стае и др.) возрастает с уве - личением численности и степени специализации вида; межвидовая борьба (взаимоотношения могут быть безразличными, вред - ными или полезными) формируются на базе пищевых отношений
между видами, а также в конкуренции за места обитания, раз |
- |
множения и т.д. Крайним выражением межвидовых отношений |
|
является межвидовая борьба, когда одна форма вытесняет другую или ограничивает ее численность на определенной территории;
борьба с неблагоприятными условиями возникает в зависимости от климатических (температура, влажность, освещенность и т.д.) или почвенных условий, влияющих на жизнедеятельность орга - низма.
В результате борьбы за существование происходит элиминация (физическая гибель или устранение при размножении) особей, которые по признакам наименее соответствуют условиям среды обитания. Таким образом, следствием борьбы за существование является естественный отбор.
Естественный отбор, по Дарвину, - это совокупность происхо - дящих в природе событий, обеспечивающих выживание наиболее
приспособленных, и преимущественное оставление ими потом |
- |
ства. Следует отметить, что естественный отбор не отбирает са |
- |
мых приспособленных, они просто сохраняются в результате |
|
элиминации менее приспособленных. В результате этого процес - са любая сохранившаяся организация, структура или функция со - ответствует состоянию приспособленности друг к другу и к окружающей среде, т.е. оказывается биологически целесообраз - ной.
Дарвин впервые на основе теории естественного отбора дал материалистическое толкование органической целесообразности, показал ее относительный характер и раскрыл пути выработки адаптаций. Он показал, что приспособленность видов на основе отбора не может быть абсолютной, она всегда относительна и адекватна лишь тем условиям среды, в которых виды длительное
132D
время существуют. Приспособления рыб целесообразны лишь в водной среде обитания и непригодны к наземной жизни; зеленая окраска саранчи является покровительственной на зеленой расти -
тельности и т.д. |
|
Дарвин считал, что возникновение новых видов происходит |
|
постепенно путем накопления полезных индивидуальных изме |
- |
нений, увеличивающихся из поколения в поколение. Чем значи |
- |
тельнее живые существа отличаются по строению и физиологи |
- |
ческим свойствам, тем большее число их групп может существо |
- |
вать на данной территории за счет ослабления борьбы за суще |
- |
ствование. С каждым поколением различия становятся все более |
|
выраженными, а промежуточные формы, сходные между собой, |
|
вымирают. Процесс видообразования, по Дарвину, происходит по принципу дивергенции, т.е. за счет расхождения признаков.
Таким образом, результатом отбора будет являться возникнове -
ние приспособлений и на этой основе – видового разнообразия. |
|
Разнообразные, меняющиеся условия среды способствуют эво |
- |
люции видов в направлении усложнения организации (млекопи |
- |
тающие, насекомые). Если виды обитают долгое время в одно |
- |
родной среде вне жесткой конкуренции, то уровень их организа |
- |
ции может оставаться на относительно низком уровне (ланцетни - ки). В постоянно меняющихся условиях среды одни виды, уменьшаясь численно, неизбежно должны погибать и уступать место другим, лучше приспособленным к новым условиям, о чем
убедительно свидетельствуют данные палеонтологии. |
|
В заключение следует подчеркнуть, что Дарвин впервые предло |
- |
жил естественно-научное объяснение эволюционного процесса. |
|
Он указал на движущие силы эволюции: наследственная измен |
- |
чивость, борьба за существование, естественный отбор; дал объ |
- |
яснение механизма видообразования и причин многообразия ви |
- |
дов, а также объяснил причины возникновения целесообразности. Признание того, что ход эволюции обусловлен естественными причинами, открыло путь к их дальнейшему научному анализу. С начала ХХ века благодаря данным науки, в первую очередь гене - тики, открылись новые возможности для анализа эволюционных
преобразований. Дальнейшее развитие идей Дарвина в работах зарубежных и отечественных исследователей: Дж. Хаксли, Дж.
133D
Симпсона, Д. Харди, Э. Майэра, Ф.Г. Добжанского, Н.В. Тимофе - ева-Ресовского, А.Н Северцова, С.С. Четверикова и ряда других – сформировало современный синтетический (основанный на дан - ных многих отраслей естествознания) этап развития теории эво - люции. Эволюционный процесс чаще всего разделяют на микро и макроэволюцию.
Микроэволюция. Критерии и структура вида. Популяция |
|
Микроэволюцией называется начальный этап эволюционных |
|
преобразований популяции: от возникновения наследственных |
|
изменений до формирования адаптации и возникновения на их |
|
основе новых видов. Изучение механизмов эволюционного про |
- |
цесса на внутривидовом уровне сделало возможным выяснить |
|
роль эволюционных факторов, сформулировать представления о |
|
эволюционных единицах (популяциях) и выявить основные мо |
- |
менты протекания эволюционных преобразований. |
|
Критерии и структура вида |
|
Видом называется совокупность особей, характеризующихся |
|
общим происхождением, наследственным сродством морфологи -
ческих, физиологических и биохимических особенностей; спо |
- |
собных скрещиваться и давать плодовитое потомство; приспо |
- |
собленных к определенным условиям среды и занимающих опре - деленный ареал.
Существует ряд общих особенностей – критериев вида. Ни один из критериев не является абсолютным, однако их комплекс явля - ется свидетельством реальности вида.
-Морфологический критерий определяет сходство внешнего и внутреннего строения, однако является недостаточно точным для различения видов, имеющих сходство в строении (виды-двойни - ки, например у комаров, тлей, крыс и др.).
-Физиологический критерий – характеризует сходство процессов жизнедеятельности особей одного вида. Он также недостаточно точен (большинство разных видов в природных условиях не скрещиваются или потомство бесплодно, однако есть исключе - ния – ряд видов канареек, зябликов, тополей и др.)
134D
- Биохимический критерий – базируется на способности синтези - ровать специфические белки, что связано с уникальными после - довательностями ДНК.
-Г енетический критерий – основывается на видовой специфич - ности набора хромосом, их индивидуальности, различиях в нук - леотидном составе молекул ДНК.
-Экологический критерий – характеризует взаимоотношения вида, его роль в биологическом круговороте веществ.
- Географический критерий определяет территорию обитания (ареал) вида.
Особи вида связаны между собой сложными взаимоотношени - ями и общностью эволюционной судьбы. Для большинства видов характерна иерархическая внутривидовая структура, состоящая из ряда внутривидовых подразделений (популяция, раса, подвид).
Популяция
Основной единицей существования, воспроизводства и эволю - ции вида является популяция. Популяция – это совокупность сво - бодно скрещивающихся особей одного вида в течение большого числа поколений, населяющих определенный ареал и частично изолированных от других популяций вида и обладающих общим генофондом. Население любого вида, как правило, распадается на относительно изолированные группы особей. Популяции форми -
руются исторически в определенных экологических условиях. |
|
Каждая популяция характеризуется экологически: популяцион |
- |
ный ареал, численность особей и ее динамика, возрастная и по |
- |
ловая структура. Эволюционируют не отдельные особи, а группы особей, объединенные в популяции. Популяцию считают про -
стейшей эволюционной единицей.
Главный фактор, определяющий единство популяции и ее от - носительную обособленность – свободное скрещивание особей. Внутри популяции каждый организм одного пола имеет равную вероятность на образование брачной пары с любым организмом другого пола. Степень свободного скрещивания особей внутри популяции гораздо выше, чем между соседними популяциям. Ор - ганизмы популяции характеризуются генетической общностью и способностью (через скрещивание) свободного обмена генетиче - ской информацией при формировании последующих поколений.
135D
Совокупность генов популяции называется |
генофондом. Важ - |
|
нейшей характеристикой генофонда являются частоты аллелей |
|
|
(генов) и генотипов. Анализ поведения генов свободно скрещи |
- |
|
вающейся популяции характеризует закон |
Харди-Вайнберга, на - |
|
званный именами ученых, впервые описавших его. Рассмотрим пример, демонстрирующий распределение пары аллельных генов
популяции: АА и аа. В первом поколении все гибриды будут ге |
- |
терозиготны (Аа). При образовании гамет, из которых формиру |
- |
ются следующие поколения, каждая половая клетка будет содер |
- |
жать один аллель. Следовательно, частота данного аллеля во вто -
ром поколении будет равна частоте половых клеток с этим алле |
- |
|||
лем. Допустим, что в гаметах аллели А и а встречаются с одина |
- |
|||
ковой частотой, равной 0,5, тогда во втором поколении частоты |
|
|||
генотипов согласно решетке Пеннета будут 0,25АА + |
0,5Аа + |
|
||
0,25аа = 1 |
|
|
|
|
гаметы |
0,5А |
|
0,5а |
|
|
|
|
|
|
0,5А |
0,25АА |
|
0,25Аа |
|
|
|
|
|
|
0,5а |
0,25Аа |
|
0,25аа |
|
|
|
|
|
|
Таким образом, частоты аллелей во втором поколении сохраня |
- |
|||
ются: 0,5А и 0,5а, значит, и в следующем поколении будут наблюдаться такие же соотношения генов и генотипов. Если частоту аллеля А обозначить через p, а частоту аллеля а – через q, то в популяции р + q = 1 соотношение генотипов в генофонде соста - вит (p + q)2 = p2 + 2pq + q2 = 1. Формулы Харди-Вайнберга дают возможность рассчитать частоту генов и генотипов в популяциях.
Закон Харди-Вайнберга показывает, что при определенных |
|
условиях частоты аллелей в популяциях из поколения в поколе |
- |
ние не меняются. В строгом смысле этот закон приложим к бес |
- |
конечно большим по численности популяциям, в которых осу |
- |
ществляется свободное скрещивание и не возникает мутаций, |
|
должен отсутствовать отбор в пользу или против какого-либо ал |
- |
леля и миграция особей с иными генотипами из соседних попу |
- |
ляций. Если в популяциях постоянно сохраняется равновесие ча - стот аллелей, то такие популяции не могут эволюционировать.
136D
Однако в природных условиях на популяции действуют внешние и внутренние факторы, нарушающие генетическое равновесие. Если популяция длительно испытывает значительное давление со стороны каких-либо внешних факторов, то неизбежно произойдет изменение генетического состава популяции. Такое длительное и направленное изменение генотипического состава популяции, ее фонда, получило название элементарного эволюционного явле - ния. Без изменения генофонда популяции эволюционный процесс невозможен.
Факторы эволюции
Изменения генотипического состава популяций происходят под действием множества событий, которые тем или иным путем в состоянии преобразовать популяции. Тем не менее возможно вы -
делить следующие основные элементарные факторы эволюции: |
|
мутационный процесс, популяционные волны, дрейф генов, изо |
- |
ляцию, естественный отбор. |
|
- Мутационный процесс |
|
Постоянная мутационная изменчивость и комбинации при |
|
скрещиваниях дают новые сочетания генов в генофонде, что не |
- |
избежно вызывает наследственные изменения в популяции. Му |
- |
тационный процесс постоянно увеличивает генетическую гетеро -
генность популяции, вследствие сохранения рецессивных мута |
- |
ций в гетерозиготах. Но сам мутационный процесс без участия |
|
других факторов эволюции не может направлять изменение при |
- |
родной популяции. Он является лишь поставщиком элементарно - го эволюционного материала.
-Популяционные волны. Дрейф генов
Популяционными волнами называют периодические колебания численности особей популяции. Причинами этих колебаний мо - гут быть различные абиотические и биотические факторы среды.
При резком сокращении численности популяции (сезонные коле -
бания, сокращение ресурсов) среди оставшихся в живых немно |
- |
гочисленных особей могут быть редкие генотипы. Если в даль |
- |
нейшем численность восстановится за счет этих особей, то это |
|
приведет к случайному изменению частот генов в генофонде. Та -
137D
ким образом, популяционные волны являются поставщиком эво - люционного материала. Если популяция мала по численности, то в результате случайных событий некоторые особи независимо от своей генетической конституции могут оставить или не оставить потомство, вследствие чего частоты некоторых генов могут резко меняться за одно поколение. Случайное изменение частот генов в генофонде популяции называется дрейф генов.
-Изоляция
Изоляция обусловлена возникновением разнообразных факто - ров, препятствующих свободному скрещиванию. Размножение идет преимущественно в пределах изолята, прекращается обмен генетической информацией с другими группами. Это способству -
ет закреплению начальной стадии изменения генофонда обосо |
- |
бившейся группы, становлению ее как самостоятельной генети |
- |
ческой системы. Различают пространственную и биологическую |
|
изоляцию. |
|
Пространственная изоляция связана с территориально-географи |
- |
ческими (водные преграды, горные хребты, места непригодные |
|
для жизни) и экологическими (расселение по разным экологиче |
- |
ским нишам) факторами разобщения популяции. Значение про |
- |
странственной изоляции зависит от величины индивидуальной |
|
активности особей вида. |
|
К биологической изоляции могут относиться особенности пове |
- |
дения, изменения строения и физиологической активности, сро |
- |
ков размножения и ряда других факторов, препятствующих скре - щиванию. После оплодотворения возможны нарушения конъюга - ции хромосом и ряд других изменений, приводящих к развитию полностью или частично стерильных гибридов, а также гибридов с пониженной жизнеспособностью.
Эволюционное значение разных форм изоляции состоит в том, что она закрепляет и усиливает генетические различия между по -
пуляциями. |
|
Изменения частот генов, вызванные рассмотренными выше фак |
- |
торами эволюции, носят случайный, ненаправленный характер и |
|
даже их совместное действие не приводит к устойчивому осу |
- |
ществлению направленного процесса эволюции. Направляющим |
|
фактором является естественный отбор. |
|
138D
-Естественный отбор
Принцип естественного отбора Дарвина имеет основополагаю - щее значение в эволюционной теории. Естественный отбор явля - ется направленным, движущим фактором эволюции органическо -
го мира. В настоящее время представления об естественном от |
- |
боре пополнены новыми фактами, расширились и углубились. |
|
Естественный отбор следует понимать как избирательное выжи |
- |
вание и возможность оставления потомства отдельными особями. Биологическое значение особи, давшей потомство, определяется вкладом ее генотипа в генофонд популяции. Отбор действует в популяции, его объектами являются фенотипы отдельных особей. Фенотип организма формируется на основе реализации информа -
ции генотипа в определенных условиях среды. |
|
Таким образом, отбор из поколения в поколение по фенотипам |
|
ведет к отбору генотипов, так как потомкам передаются не при |
- |
знаки, а генные комплексы. Для эволюции имеют значение не |
|
только генотипы, но и фенотипы и фенотипическая изменчи |
- |
вость. |
|
В процессе экспрессии ген может оказывать влияние на многие |
|
признаки. Поэтому в сферу действия отбора могут включаться не |
|
только свойства, повышающие вероятность оставления потом |
- |
ства, но и признаки, которые не имеют прямого отношения к вос - производству. Они отбираются опосредованно в результате кор - реляции. В ряде случаев отбор может быть направлен на создание взаимоприспособлений видов друг к другу (цветки растений и посещающие их насекомые). Так же могут создаваться признаки, вредные для отдельной особи, но обеспечивающие выживание вида в целом (ужалившая пчела гибнет, но, нападая на врага, она сохраняет семью). В целом отбор играет творческую роль в при - роде, поскольку из ненаправленных наследственных изменений закрепляются те, которые могут привести к образованию новых групп особей, более совершенных в данных условиях.
Различают три основные формы отбора: стабилизирующий, дви - жущий и разрывающий (дизруптивный).
139D
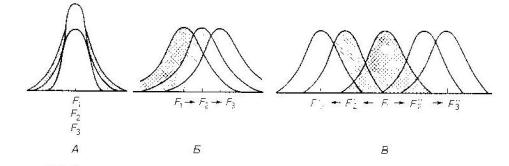
А – стабилизирующий отбор Б – движущий отбор В – дизруптивный отбор
F1-F3 – поколения (заштрихованы варианты, устраняемые отбо - ром)
1). Стабилизирующий отбор способствует сохранению призна -
ков вида в относительно постоянных условиях среды. Он под |
- |
держивает средние значения, выбраковывая мутационные откло |
- |
нения ранее сформировавшейся нормы. Стабилизирующая форма отбора действует до тех пор, пока сохраняются условия, повлек - шие образование того или иного признака. Примером стабилизи - рующего отбора являются наблюдения за избирательной гибелью домовых воробьев при неблагоприятных погодных условиях. У выживших птиц различные признаки оказались близки к средним значениям, а среди погибших сильно варьировались. Примером действия стабилизирующего отбора в популяции людей является большая выживаемость детей со средней массой тела.
2).Движущий отбор благоприятствует изменению среднего значения признака в измененных условиях среды. Он обусловли - вает постоянное преобразование приспособлений видов соответ - ственно изменениям условий существования. Особи популяции имеют некоторые отличия по фенотипу и генотипу. При длитель -
ном изменении внешней среды преимущественно в жизнедея |
- |
тельности и размножении может получить часть особей вида с |
|
некоторыми отклонениями от средней нормы. Это приведет к из -
менению генетической структуры, возникновению эволюционно |
|
новых приспособлений и перестройке организации вида. Вариа |
- |
ционная кривая смещается в направлении приспособления к но |
- |
вым условиям существования. |
|
140D |
|