

-
Автономная саморегуляция м/о (дыхательный контроль). Энергетический заряд клетки, возможный диапазон его значений.
АВТОНОМНАЯ САМОРЕГУЛЯЦИЯ ЦТК
В ЦТК два ключевых фермента:
1) цитратсинтаза (1-я реакция)
2) изоцитратдегидрогеназа (3-я реакция)
Оба фермента аллостерически ингибируются избытком АТФ и НАДН2. Изоцитратдегидрогеназа сильно активируется АДФ. Если АДФ нет, то этот фермент неактивен. В условиях энергетического покоя концентрация АТФ увеличивается, и скорость реакций ЦТК мала - синтез АТФ уменьшается.
Изоцитратдегидрогеназа ингибируется АТФ намного сильнее, чем цитратсинтаза, поэтому в условиях энергетического покоя повышается концентрация цитрата, и он выходит в цитоплазму по градиенту концентраций путем облегченной диффузии. В цитоплазме цитрат превращается в Ацетил-КоА, который участвует в синтезе жирных кислот.
ЭНЕРГЕТИЧЕСКИЙ ЗАРЯД КЛЕТКИ (ЭЗК)
Многие процессы метаболизма регулируются соотношением АТФ/АДФ. Это соотношение характеризует энергетический статус (состояние) клетки (Энергетический Заряд Клетки).
[АТФ]+ 1/2[АДФ]
ЭЗК = ------------------------
[АТФ] + [АДФ] + [АМФ]
Теоретически ЭЗК может меняться от 0 до 1. Обычно в норме величина ЭЗК поддерживается в клетке в пределах 0.85-0.90.
ОБРАТИТЕ ВНИМАНИЕ! Метаболические пути, приводящие к образованию АТФ, ингибируются высоким ЭЗК; метаболические пути, ведущие к использованию АТФ, активируются высоким ЭЗК.
2. Написать реакции цтк от суккцинил-КоА до оксалоацетата.Энерг итог.
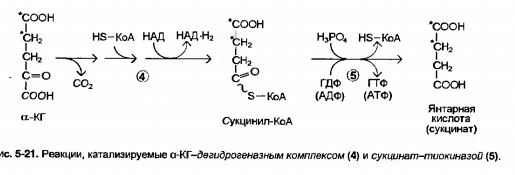
Сукцинат 1,5 АТФ
альфа-кетоглутарат 2,5
итог
лллллллллллллл
-
Окислительные реакции, катализ оксидазами. Строение и роль.



4. Активные формы кислорода, их свойства и био знаечние.
Включают ионы кислорода, свободные радикалы и перекиси как неорганического, так и органического происхождения. Это, как правило, небольшие молекулы с исключительной реактивностью благодаря наличию неспаренного электрона на внешнем электронном уровне.
Главным источником АФК в клетках являются митохондрии. Обычно примерно 98% всего кислорода, поступающего в клетки, используется для окисления субстратов с образованием АТФ и выделением тепла, и лишь 2% используется в реакциях образования АФК, которое может значительно возрастать при усиленном поступлении кислорода в клетки или нарушении работы электронно–транспортной цепи митохондрий.
Молекулярный кислород не токсичен для клеток, однако опасность представляют продукты его неполного окисления: перекисные соединения, супероксидные радикалы, синглетный кислород и др. В связи с биологической активностью эти соединения получили название активные формы кислорода (АФК). Появление АФК вызвано тем, что молекулярный кислород (02) может перехватывать электроны у некоторых переносчиков цепи электронного транспорта. В результате одноэлектронного восстановления молекулы кислорода образуется супероксидный радикал или анион-радикал:
02 + е -> 02-.
Образование АФК происходит и при взаимодействии озона с кислородом:
03 + 02 -> 202 + 02-.
Супероксидный радикал — заряженная частица, окруженная молекулами воды. Поэтому 02- не может преодолеть мембрану, оказывается «запертым» в клетке и становится источником других форм АФК, например перекиси водорода:
02- + е + 2Н+ —> Н202.
Перекись водорода, в свою очередь, восстанавливается и дает гидроксил-радикал:
Н202 + е + Н+ -> ОН-.
Реакционная способность последнего чрезвычайно высока, поэтому гидроксил-радикал способен окислить практически любое вещество клетки, включая ДНК. Концентрации АФК в тканях невысоки и составляют 10-8—10-11 М. АФК вызывают образование органических гидропероксидов (ROOH) ДНК, белков, липидов. Этот процесс называют перекисным окислением. Гидропероксиды в ходе метаболизма способны превращаться в различные окисленные соединения — спирты, альдегиды и др. Так, в ходе перекисного окисления липидов (ПОЛ) снижается содержание ненасыщенных жирных кислот, образуются различные производные жирных кислот, а затем такие метаболиты как малоновый диальдегид, этан и др. АФК образуются в различных частях клетки. У животной клетки наибольший вклад вносит дыхательная цепь митохондрий. У растений эти процессы происходят еще и в хлоропластах. Поэтому у растений возможность образования АФК выше, чем в животной клетке.
В тканях аэробных организмов в процессе метаболизма постоянно образуются продукты неполного восстановления кислорода. Активные формы кислорода и радикалы синтез в организме, выполняют не только вредные, но и множество полезных для клетки функций. Так, образование супероксид-аниона и гипохлорита клетками иммунной системы используется организмом при защите от инфекций и других чужеродных факторов. Для некоторых тканей, в частности, для мозга, характерен повышенный синтез простагландинов, тромбоксанов и лейкотриенов. Этот процесс требует участия супероксид-аниона, взаимодействующего с другим компонентом этой системы, арахидоновой кислотой - соединением, высвобождающимся из мембранных фосфолипидов в ходе индуцируемого АФК перекисного окисления липидов.
АФК активно участвуют в процессах передачи клеточного сигнала. Так, например, свободные радикалы, которые образуются в цитозоле клетки в ответ на стимуляцию факторами роста, участвуют в регуляции пролиферативного. Имеется много публикаций, свидетельствующих, что активные формы кислорода стимулируют деление различных типов клеток.
Механизм, по которому активные формы кислорода участвуют в передаче митотического сигнала, неизвестен. Возможно, что АФК могут активировать такие ферменты, как митоген-активируемую киназу p38, что, в свою очередь, приводит к активации транскрипционного фактора HIF-1 и экспрессии соответствующих генов. Показано, что они включают каскад реакций, которые передают митотический сигнал при воздействии "физиологических" агентов, таких как факторы роста: активируют в клетке различные транскрипционные факторы, но наиболее эффективно - продукт гена c-jun и ядерный транскрипционный фактор NF-kB. Свободные радикалы обладают активностью, модулирующей транскрипционные факторы, как NF-kB и активирующий белок-1 (AP-1). В случае NF-kB показано, что он становится транскрипционно активным после того, как происходит опосредованная АФК деградация IkB белка, ингибирующего NF-kB. Также показана роль АФК в качестве вторичных мессенджеров в сигнальных каскадах, запускаемых такими факторами как TGF-b1, PDGF, ATII, FGF-2 и эндотелин.
Еще одной мишенью АФК может являться Na/K-АТФаза, белок, отвечающий за электрогенный транспорт ионов калия и натрия через клеточную мембрану. В нормальных условиях нейрональная Na/K-АТФаза потребляет от 15% до 40% энергии клетки, направляя ее на поддержание ионных градиентов. Избыточная активация глутаматных рецепторов и повышение уровня АФК приводит к обратимому ингибированию фермента. Известно разнонаправленное влияние глутаматных рецепторов первой и третьей групп на активность Na/K-АТФазы. Так, АФК усиливают ингибирующее влияние NMDA-рецепторов на активность Na/K-АТФазы, а mGluI предотвращают ингибирование ее NMDA — рецепторами.