

гиста / Эмбриология, тератология и основы репродукции человека _ В.Н. Запорожан, В.К. Напханюк, Е.Л. Холодкова. - О._ ОГМУ, 2000. - 378 с
.pdfГЛАВА 1
ОБЩАЯ ЭМБРИОЛОГИЯ
1.1. ВВЕДЕНИЕ
Эмбриология человека — наука о развитии зародыша от момента оплодотворения до периода, когда его организм приобретает основные морфологические черты взрослой особи и способность к самостоятельной жизнедеятельности.
Процессы закладки, формирования и развития тканей и органов контролируются геномом, который действует на нескольких уровнях организации. Результаты экспериментальных исследований на эмбрионах животных помогли понять процессы развития человека. Широко внедряются в практику и теорию данные о клеточных и субклеточных структурах, исследованных различными методами молекулярной биологии, клеточной биологии, биохимии и генетики.
Эмбриология человека как наука в последнее время имеет исключительно важное значение, поскольку позволяет установить механизмы влияния различных эндогенных и экзогенных факторов на закладку, развитие и созревание половых клеток, образование тканей, органов и систем органов, на процессы, контролирующие репродуктивную функцию и обеспечивающие поддержку гомеостаза зародыша человека. Кроме того, эмбриология играет ведущую роль в изучении и предопределении развития уродств и аномалий развития, а также сохранении здорового генофонда государства.
Эмбриология формировалась в тесной взаимосвязи с развитием медицинской науки вообще. По мере совершенствования методов исследования возникали различные теории, которые пытались объяснить те или иные процессы, происходящие в
9
эмбриональном периоде. Еще в XVIII веке было установлено, что свойства организма заложены в сперматозоидах или в ооцитах. Иначе говоря, формирование организма заранее предопределено (теория преформационизма). В 1759 г. Caspar Friedrich Wolff предположил, что организмы развиваются стадийно, с постепенным видимым усложнением их строения. Другими словами, дифференцировка так же необходима, как и рост, так как разнородность формируется из однородности. В настоящее время предложена теория эпигенеза, по которой развитие организма заключается в успешном, не предопределенном формировании его частей. Однако, тот факт, что наследственная информация хранится и реализуется при помощи генома, иногда выглядит как современная преформационистская концепция.
В XIX веке пользовалась большой популярностью теория, названная биогенетическим законом Геккеля—Мюллера. Суть ее состояла в том, что индивидуальное развитие (онтогенез) сходно (“повторяет”) со стадиями эволюции предков (филогенезом). Теория повторения, тем не менее, по словам de Beer, имела “прискорбное влияние на развитие эмбриологии”. Исследования Carl Ernst von Baer в 1828 г. были намного ближе к истине. В соответствии с “законами” von Baer, общие структуры (мозг, нотохорда) формируются в процессе развития раньше, чем специальные (конечности, волосы). В дальнейшем развитие зародыша человека все больше и больше отклоняется от развития других видов животных. Конечно, поздние этапы развития человека отличаются от таковых у животных, однако на ранних стадиях формирования они имеют определенное сходство. Фарингеальные карманы позвоночных, например, не являются ни жабрами, ни складками, хотя у рыб эти области превращаются в жаберные щели, у птиц, рептилий и млекопитающих — в гланды и тимус.
Согласно современным представлениям формирование зародыша человека происходит в результате процессов индукции, детерминации, роста, размножения, миграции клеток, межклеточных взаимодействий и клеточной смерти.
Индукция — это процесс влияния одних тканей или участков зародыша на другие, вследствие чего происходит стимуляция дифференцирования их в определенном направлении. Примером индукции может быть влияние глазного пузырька
10
на формирование хрусталика из подлежащей эктодермы. Определенный участок эмбриона, который вызывает образование и определяет дальнейшее развитие других структур, называется индуктором. Так, первичный индуктор образования позвоночной оси Hans Spemann назвал организатором. В 20-х годах XX века он проводил эксперименты на амфибиях с трансплантацией хордомезодермального участка и обнаружил образование вторичной оси тела. Многие ученые продолжили изучение процессов индукции. Было установлено, что гены, содержащие гомеобоксы (участки ДНК, кодирующие синтез аминокислот), могут принимать участие в регуляции функциональной способности организатора.
В первичной индукции полипотентные клетки-мишени определенного участка эктодермы “запрограммированы” к развитию в нервную ткань. Вторично детерминируется формирование определенной структуры нервной системы. Сигнальные частицы (диффундирующие молекулы), которые переносят информацию от индуктора к мишени, неизвестны.
Детерминация — это процесс, в результате которого происходит определение дальнейшего пути развития клеток на генетической основе вследствие блокирования отдельных компонентов генома. Процесс детерминации является основой дифференциации, которая представляет собой процесс развития специализированных клеток из однородных (индифферентных). Различают три основных вида дифференциации: оотипическую, когда исходный материал представлен презумптивными участками цитоплазмы зиготы; бластомерную — в период бластоцисты; зачаточную, характеризующуюся появлением в пределах одного зародышевого листка различных тканей.
Процесс размножения клеток — это увеличение их количества, обусловленное аутосинтетической и гетеросинтетической деятельностью клеток. Аутосинтетическая деятельность направлена на усиление процессов репродукции, тогда как гетеросинтетическая — на формирование специфических структур или синтез и выделение специфических продуктов. Если между гетеро- и аутосинтетической деятельностью клеток существуют ярко выраженные антогонистические отношения, то у тканевых систем имеется два способа соединения процессов репродукции и дифференциации: 1) все репродуктивные процессы сконцентрированы в период эмбриогенеза, когда проис-
11
ходит формирование основного запаса клеток (нервная ткань); 2) образование запасов малодифференцированных пролиферирующих клеток, которые обеспечивают возникновение новых (кроветворная ткань). По этому признаку все ткани можно подразделить на три типа: 1) стационарные — возраст образующих их клеток соответствует возрасту организма (нейроциты); 2) восстанавливающиеся — количественное постоянство клеточного состава достигается за счет сбалансированного соотношения погибших и образованных клеток; 3) растущие — характеризуются постоянным ростом клеток.
Миграция клеток — это активное массовое морфогенетическое перемещение клеток из одной части зародыша в другую, в результате которого образуются ткани и органы.
Рост клеток — процесс развития и организации клеток вследствие ряда сложных преобразований, происходящих от момента формирования клетки до последующего деления.
Ведущее значение для прогрессирующей дифференциации клеток эмбриональных зачатков имеют процессы взаимодействия между клетками разных зачатков или однотипными клетками одного и того же зачатка. Установлено, что для нормального развития и существования эпителия необходим постоянный или временный контакт с развивающейся соединительной тканью.
Клеточная смерть — это необратимая остановка всех функций клетки и ее связей с окружающей средой. Запрограммированная клеточная смерть (апоптоз) контролируется генетически и является нормальной особенностью эмбрионального развития, играя важную роль в морфогенезе. Например, смерть межпальцевых клеток способствует формированию формы конечностей и отделению пальцев от ладонной и стопной пластинок. Другой пример — установление окончательного количества нейронов в двигательных столбах спинного мозга и в чувствительных ганглиях. Во многих случаях объем клетки уменьшается, хроматин конденсируется, клетка умирает, фрагментируется и фагоцитируется окружающими клетками. Некоторые клетки, однако, разрушаются своими собственными лизосомами (аутофагия), а другие, по-видимому, без участия лизосом. В патологических условиях клеточная смерть (некроз) характеризуется набуханием клеток, вследствие чего клеточная оболочка разрывается.
12

1.2. ПРОГЕНЕЗ И ОПЛОДОТВОРЕНИЕ
Рассмотрим процессы образования гамет (сперматозоидов и яйцеклеток), выхода ооцитов (овуляция) и оплодотворения.
Гаметогенез
Под гаметогенезом понимают образование половых клеток (гамет), т. е. сперматозоидов и ооцитов. Эти клетки образуются в гонадах — половых железах — яичках и яичниках. Рост гамет происходит в результате последовательных делений определенной линии клеток (половые клетки) и клеток, не связанных напрямую с образованием половых клеток (соматические клетки). Во время дифференциации гамет диплоидные клетки (с двойным набором хромосом как у соматических клеток) называются первичными, а гаплоидные (с одинарным набором хромосом) — вторичными. Редукция числа хромосом от 46 (диплоидный или 2 n) до 23 (гаплоидный или 1 n) достигается во время клеточного деления — мейоза. Мейоз характеризуется двумя последовательными делениями, во время которых происходит образование пар гомологичных хромосом, обмен генетическим материалом и формирование новых хромосомных комбинаций. Из 46 хромосом — 44 представляют собой аутосомы и 2 — половые хромосомы: X и Y. Мужской пол определяется XY набором половых хромосом, а женский — XX.
Процессы спаривания и обмена ограничены небольшими участками X- и Y-хромосом. Большая часть Y-хромосомы генетически инертна, лишь небольшой отрезок содержит SRY (область, определяющая пол) ген, который обусловливает развитие по мужскому типу, инициируя формирование яичек, а не яичников.
В соответствии с гипотезой о неактивной X-хромосоме, во время раннего развития млекопитающих женского пола одна из двух X-хромосом становится генетически инертной, транскрипционно неактивной, цитологически гетерохроматичной, появляющаяся как глыбка полового хроматина в соматических клетках. Более того, согласно гипотезе о единственно активной X-хромосоме (в клетках с несколькими X-хромосома- ми) только одна будет активной. Реактивация X-хромосомы происходит в половых клетках, вероятно, в конце мейоза. Обе X-хромосомы функционируют в периоде роста и созревания ооцита.
13
Примордиальные половые клетки (ППК), или гоноциты,
плохо распознаваемы у ранних человеческих эмбрионов. Доказано, что они образуются на стадии бластоцисты. Полагают, что они обособляются не позднее 2-й недели, а возможно, и намного раньше. В 31/2 недели они могут обнаруживаться в желточном мешке и первичной кишке. Неделей или двумя позже они мигрируют в гонады.
Сперматогенез
Сперматогенез — образование сперматозоидов на протяжении всей жизни человека — с пубертатного периода до глубокой старости. Этот процесс происходит в яичках, являющихся также эндокринной железой вследствие продукции интерстициальными клетками тестостерона. До полового созревания сперматогонии в семенных канальцах яичка остаются относительно неактивными. После полового созревания происходит стимуляция интерстициальных клеток, сперматогонии пролиферируют (сперматоцитогенез) и некоторые из них превращаются в первичные сперматоциты. В результате первого деления созревания (мейоз I) образуются сперматиды, которые затем преобразуются в сперматозоиды (спермиогенез). Формирование сперматозоида из сперматогонии происходит в течение 9-и недель. Сперматозоид состоит из головки (с акросомным колпачком), жгутика или хвоста, который имеет шейку, промежуточную и конечную части.
Сперматозоид достигает внешней среды, проходя сложный путь: извитые семенные канальцы, сеть яичка, выносящие канальцы яичка, дольки придатка яичка, проток придатка, семявыносящий проток, семявыбрасывающий проток, простатическую, перепончатую и губчатую части мочеиспускательного канала. Семенная жидкость кроме сперматозоидов, содержит секреты придатка, семенных пузырьков, простаты, бульбоуретральных и уретральных желез.
Сперматозоиды созревают в придатке яичка. Попадая в женские половые пути, они сохраняют способность к оплодотворению не более 72 часов, что зависит от внутренних и внешних (маточных, трубных, окружающих ооцит) факторов. Этот процесс называется капацитацией и включает в себя удаление с поверхности сперматозоида определенных макромолекул (главным образом протеинов и гликопротеинов семенной жид-
14
кости). Капацитация, способствующая повышению двигательной активности сперматозоида, является необходимым приготовлением к акросомальной реакции. Последняя стимулируется массивным вхождением внеклеточного кальция внутрь сперматозоида. В результате этой реакции, которая происходит на поверхности прозрачной зоны, высвобождаются ферменты (акрозин, гиалуронидаза), что вызывает пенетрацию зоны. Образуется несколько точек слияния между плазмолеммой и подлежащей ей наружной акросомальной мембраной, в результате образуются щели, через которые проникает акросомальное содержимое. Данная реакция необходима для прохождения сперматозоидом прозрачной зоны. Все эти процессы достаточно сложны и еще окончательно не изучены. Исследования в этом направлении продолжаются.
Оогенез
Оогенез представляет собой процесс образования и созревания женских половых клеток. Женские гаметы образуются из оогоний, которые, в свою очередь, являются производными примордиальных половых клеток. Оогонии размножаются митотически и превращаются в первичные ооциты. К середине эмбрионального развития число оогоний достигает 7 млн, после чего их количество уменьшается и к моменту рождения составляет приблизительно 2 млн. Из них образуется несколько тысяч ооцитов, из которых несколько сотен достигает зрелости и освобождается (овулирует) в течение репродуктивного периода женщины, продолжающегося около 30 лет. Профаза первого мейоза начинается еще в эмбриональном периоде, но “замирает” на стадии диплотены, которая длится весь период детства. Фолликулярные клетки (зернистый слой) играют важную роль в блокировании завершения мейоза.
После полового созревания первый мейоз заканчивается, образуется вторичный ооцит (размером 120 мкм и более) и первое направительное тельце, которое представляет собой ооцит с крайне малым объемом цитоплазмы. Во вторичном ооците завершается первое мейотическое деление и начинается второе. Процесс превращения оогонии во вторичный ооцит длится от 12 до 50 лет. Второй мейоз заканчивается после выхода фолликула (овуляции), но только после проникновения сперматозоида в яйцеклетку (рис. 1.2.1).
15
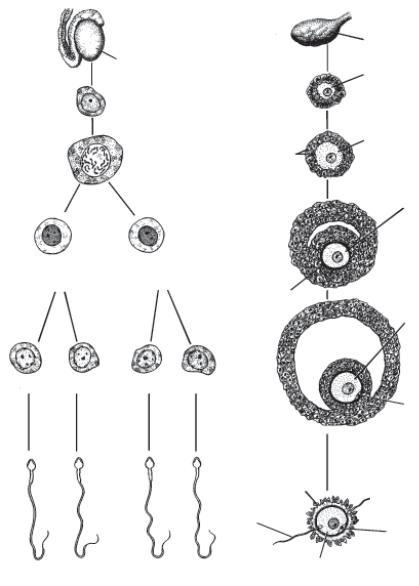
|
|
|
|
|
ГАМЕТОГЕНЕЗ |
|
|
|
|
|
СПЕРМАТОГЕНЕЗ |
|
|
ООГЕНЕЗ |
|
|
|
||||
|
|
|
|
|
|
|
|
яичник |
||
|
|
|
яичко |
|
|
|
первичный |
|||
|
|
|
|
|
|
|
|
ооцит 46,XX |
||
|
|
|
сперматогония |
|
|
в первичном |
||||
|
|
|
|
|
|
фолликуле |
||||
|
|
|
46,XY |
|
|
|
||||
|
|
|
|
|
|
|
|
|||
|
|
|
первичный |
фоллику- |
|
|
первичный |
|||
|
|
|
|
ооцит 46,XX |
||||||
|
|
|
сперматоцит |
лярные |
|
|
в растущем |
|||
|
|
|
46,XY |
клетки |
|
|
фолликуле |
|||
|
|
|
|
|
первое |
|
|
|
|
|
|
|
|
мейотическое |
|
1 |
|||||
|
|
|
|
|
деление |
|
|
|
|
|
|
23,X |
23,Y |
|
|
|
|
первое |
|||
вторичные сперматоциты |
прозрачная |
|
|
|||||||
|
мейотическое |
|||||||||
|
|
|
|
|
|
|
||||
|
|
второе мей- |
|
оболочка |
|
|
деление |
|||
|
|
|
|
|
2 |
|||||
|
|
отическое |
|
|
|
|||||
|
|
деление |
|
|
полость |
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
23,X |
23,X |
23,Y |
23,Y |
|
|
|
|
|
||
|
|
сперматиды |
|
|
|
|
первое |
|||
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
полярное |
|||||
|
|
СПЕРМИОГЕНЕЗ |
|
|
|
|
||||
|
|
|
|
|
|
лучистый |
|
тельце |
||
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
второе |
|||
|
|
|
|
|
|
венец |
|
|
||
|
|
|
|
|
|
|
|
мейотическое |
||
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
сперма- |
|
|
деление |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
тозоид |
|
|
|
второе |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
полярное |
|
|
|
|
|
|
|
|
|
|
|
тельце |
|
|
|
|
|
|
|
|
|
|
23,X |
23,X |
23,X |
23,Y |
23,Y |
зрелый |
|
|
|
|||
|
|
|
|
|
|
|
|
|
||
ооцит
16

Яичниковый фолликул
Яйцеклетка находится в пузырьке, который называется яичниковый фолликул. Образование фолликулов начинается гдето в середине пренатальной жизни. Предполагается, что фолликулярные клетки происходят как из целомического эпителия, так и из мезонефральных клеток. Вначале первичный ооцит окружен одним слоем плоских фолликулярных клеток (примордиальный фолликул), затем — слоем кубических или цилиндрических клеток (первичный фолликул), в последующем — многослойным эпителием (вторичный фолликул) и, наконец, клетками, образующими полость, или антрум, заполненную жидкостью (третичный или пузырчатый фолликул). В это время у ооцита появляется полупрозрачная оболочка, которую, предположительно, образуют ооцит и фолликулярные клетки. Эта зона состоит из гликопротеинов.
Исследователи считают, что непосредственно перед овуляцией фолликул, в котором теперь находится вторичная яйцеклетка, становится зрелым и достигает 2–3 см в диаметре. Цитоплазматические отростки внутреннего слоя фолликулярных клеток проникают в прозрачную зону, но перед овуляцией отходят. Ооцит считается зрелым, когда достигает метафазы второго деления меойза. Обычно у человека только один фолликул достигает зрелости в каждом овариальном цикле, а большинство вторичных и третичных фолликулов подвергаются обратному развитию, образуя атретические фолликулы. Атрезия — возможность уменьшения числа ооцитов с возрастом.
По мере образования полости, фолликулярные клетки смещаются к периферии и формируют зернистый слой, кнаружи от которого строма яичника уплотняется и образует железистый слой — внутреннюю теку. Участок, где находится ооцит,
Рис. 1.2.1. Сравнительные схемы сперматогенеза и оогенеза. На рисунке не показаны оогонии, так как они дифференцируются в первичные ооциты еще до рождения. На каждой стадии указан хромосомный состав. Следует отметить, что 1) вследствие мейотических делений диплоидный набор хромосом становится гаплоидным; 2) из одного первичного сперматоцита развивается четыре зрелых сперматозоида, тогда как из одного первичного ооцита развивается только один зрелый ооцит; 3) в период оогенеза цитоплазма накапливается для формирования одной крупной клетки — зрелого ооцита.
1 — первичный ооцит 46,XX в большем фолликуле; 2 — вторичный ооцит 23,Х в зрелом фолликуле
17
окруженный фолликулярными клетками, называют яйценосным бугорком, который выступает в полость фолликула. Окончательное развитие фолликула и овуляция зависят от стимуляции яичника фолликулостимулирующим (ФСГ) и лютеинизирующим (ЛГ) гормонами гипофиза. Сам яичниковый фолликул также является источником образования гормонов, в основном эстрогенов.
Овуляция
Овуляция — выход вторичного ооцита из зрелого яичникового фолликула. Она вызывается высвобождением ЛГ из гипофиза. Простагландины также вовлечены в этот процесс. В результате овуляции ооцит попадает в новое окружение (маточная труба), где может произойти оплодотворение. Овуляции начинаются в пубертатном периоде и прекращаются в менопаузе. Вследствие циклических процессов это повторяется приблизительно каждые 28 дней, но может и значительно варьировать (21–35 дней).
Ооплазма вторичного ооцита окружена слоем гликопротеинов, прозрачной оболочкой и небольшим количеством фолликулярных клеток яйценосного бугорка. Эти клетки продолжают удерживаться вокруг ооцита после овуляции, образуя лучистый венец. В подоболочечном пространстве находится полярное тельце. Освобожденная яйцеклетка, хотя формально и находится в брюшной полости, оказывается тесно прижатой к абдоминальному отделу маточной трубы, в норме попадает в нее и продвигается к матке. Яйцеклетка считается способной к оплодотворению в течение 24-х часов нахождения в женских репродуктивных органах.
В “идеализированном” 28-дневном цикле время овуляции — приходится на 14±1 день до предполагаемого начала следующей менструации. Наиболее вероятными являются дни в промежутке между 9-м и 18-м днями цикла.
Выход яйцеклетки из маточной трубы или наступление беременности означают, что овуляции происходят. Косвенными методами определения наличия овуляции являются: биопсия эндометрия (наличие секреторного эндометрия), влагалищный мазок (повышение ороговения эпителия), электрическое сопротивление влагалища, ультразвук (увеличение размера фолликула), шеечная слизь (прозрачная и тягучая перед овуляцией:
18