

3. Эндокринная функция поджелудочной железы
Эндокринная часть поджелудочной железы образована лежащими между ацинусов панкреатическими островками, или островками Лангерганса - светлыми образованиями размером 50— 500 мкм, состоящими из разных типов эпителиальных клеток (альфа-, бета-и дельта-клеток). Островки составляют около 1,5% объема железы; у человека их насчитывается около 1 млн.
Островки состоят из клеток – инсулоцитов, среди которых на основании наличия в них различных по физико-химическим и морфологическим свойствам гранул выделяют 5 основных видов:
- бета-клетки, синтезирующие инсулин (60—80 %);
- альфа-клетки, продуцирующие глюкагон (20 %);
- дельта-клетки, образующие соматостатин;
- D1-клетки, выделяющие ВИП;
 -
PP-клетки, вырабатывающие панкреатический
полипептид.
-
PP-клетки, вырабатывающие панкреатический
полипептид.
Кроме того, методами иммуноцитохимии и электронной микроскопии было показано наличие в островках незначительного количества клеток, содержащих гастрин, тиролиберин и соматолиберин.
Островки представляют собой компактные пронизанные густой сетью фенестрированных капилляров скопления упорядоченных в гроздья или тяжи внутрисекреторных клеток. Клетки слоями окружают капилляры островков, находясь в тесном контакте с сосудами; большинство эндокриноцитов контактируют с сосудами либо посредством цитплазматических отростков, либо примыкая к ним непосредственно.
3.1. Инсулин: химическая природа, регуляция секреции, механизм действия, роль в обмене.
Инсулин представляет собой простой полипептид с молекулярной массой около 6000, состоящий из двух линейных цепей: А — с 21 аминокислотой и В — с 30 аминокислотами, соединенных двумя бисульфидными мостиками. В цепи А есть еще один «внешний» бисульфидный мостик. Структура инсулина у различных животных в основном совпадает. В настоящее время принята точка зрения, согласно которой вначале синтезируется одна цепь аминокислот с последующим свертыванием молекулы и образованием бисульфидных связей. В этом процессе участвует так называемый соединительный пептид, или пептид С, состоящий из 33 аминокислот и соединяющий цепи А и В. После свертывания образуется предшественник инсулина, или проинсулин, с молекулярной массой свыше 9000.
Биологическая активность проинсулина составляет примерно 10% активности инсулина. Соединительный пептид С отщепляется от инсулина в гранулах β-клеток перед секрецией в кровь и поступает в нее в количестве, эквимолекулярном инсулину. Определение уровня пептида С является более надежным индикатором секреции β-клеток, чем определение собственно инсулина, так как последний в отличие от С-пептида частично задерживается в печени, может подвергаться воздействию циркулирующих противоинсулиновых антител и подвергаться другим воздействиям, искажающим истинную картину. Разъединение цепей А и В приводит к полной инактивации инсулина.



Регуляция секреции инсулина происходит на основе простой обратной связи, регулирующим фактором в которой является уровень гликемии: секреция инсулина повышается при гипергликемии и понижается при гипогликемии. При этом (3-клетки являются одновременно хеморецепторами и эффекторами. К основной регуляции функции (3-клеток присоединяется модулирующее влияние некоторых гормональных факторов: СТГ, гормонов надпочечников — стероидов и катехоламинов. Вероятно, в интраинсулярной регуляции участвуют глюкагон и соматостатин, а также ионы кальция, связанные с кальмодулином. Определенное влияние на секрецию инсулина могут оказывать и нервные факторы (блуждающий нерв). Косвенное участие в регуляции секреции инсулина принимают адреналин, глюкагон, глюкокортиконцы и тиреоидные гормоны, а соответственно, АКТГ и ТТГ, вызывающие гипергликемию. Предполагается, что существуют также интраинсулярные регуляторные связи между α-, β- и γ-клетками, в основе которых лежит непосредственное (не связанное с гипергликемией) стимулирующее действие глюкагона на секрецию инсулина. К другим эндогенным факторам, оказывающим стимулирующее воздействие на секрецию инсулина, относятся некоторые аминокислоты (особенно лейцин и аргинин), ионы натрия, холинергические вещества; высокая внутриклеточная концентрация калия блокирует секрецию инсулина.
Секреция инсулина происходит постоянно, и около 50% инсулина, высвобождаемого из β-клеток, никак не связано с приемом пищи или иными влияниями. В течение суток поджелудочная железа выделяет примерно 1/5 от запасов имеющегося в ней инсулина.
Главным стимулятором секреции инсулина является повышение концентрации глюкозы в крови выше 5,5 ммоль/л, максимума секреция достигает при 17-28 ммоль/л. Особенностью этой стимуляции является двухфазное усиление секреции инсулина:
первая фаза длится 5-10 минут и концентрация гормона может 10-кратно возрастать, после чего его количество понижается,
вторая фаза начинается примерно через 15 минут от начала гипергликемии и продолжается на протяжении всего ее периода, приводя к увеличению уровня гормона в 15-25 раз.
Чем дольше в крови сохраняется высокая концентрация глюкозы, тем большее число β-клеток подключается к секреции инсулина.
Стимуляция синтеза инсулина происходит от момента проникновения глюкозы в клетку до трансляции инсулиновой мРНК. Она регулируется повышением транскрипции гена инсулина, повышением стабильности инсулиновой мРНК и увеличением трансляции инсулиновой мРНК.
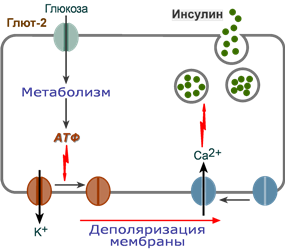
Стимуляция секреции инсулина
1. После проникновения глюкозы в β-клетки (через ГлюТ-1 и ГлюТ-2) она фосфорилируется гексокиназой IV (глюкокиназа, обладает низким сродством к глюкозе),
2. Далее глюкоза аэробно окисляется, при этом скорость окисления глюкозы линейно зависит от ее количества,
3. В результате нарабатывается АТФ, количество которого также прямо зависит от концентрации глюкозы в крови,
4. Накопление АТФ стимулирует закрытие ионных K+-каналов, что приводит к деполяризации мембраны,
5. Деполяризация мембраны приводит к открытию потенциал-зависимых Ca2+-каналов и притоку ионов Ca2+ в клетку,
6. Поступающие ионы Ca2+ активируют фосфолипазу C и запускают кальций-фосфолипидный механизм проведения сигнала с образованием ДАГ и инозитол-трифосфата,
7. Появление инозитол-трифосфата в цитозоле открывает Ca2+-каналы в эндоплазматической сети, что ускоряет накопление ионов Ca2+ в цитозоле,
8. Резкое увеличение концентрации в клетке ионов Ca2+ приводит к перемещению секреторных гранул к плазматической мембране, их слиянию с ней и экзоцитозу кристаллов зрелого инсулина наружу,
9. Далее происходит распад кристаллов, отделение ионов Zn2+ и выход молекул активного инсулина в кровоток.
Описанный ведущий механизм может корректироваться в ту или иную сторону под действием ряда других факторов, таких как аминокислоты, жирные кислоты, гормоны ЖКТ и других гормоны, нервная регуляция.
Из аминокислот на секрецию гормона наиболее значительно влияют лизин и аргинин. Но сами по себе они почти не стимулируют секрецию, их эффект зависит от наличия гипергликемии, т.е. аминокислоты только потенциируют действие глюкозы.
Свободные жирные кислоты также являются факторами, стимулирующими секрецию инсулина, но тоже только в присутствии глюкозы. При гипогликемии они оказывают обратный эффект, подавляя экспрессию гена инсулина.
Логичной является положительная чувствительность секреции инсулина к действию гормонов желудочно-кишечного тракта – инкретинов (энтероглюкагона и глюкозозависимого инсулинотропного полипептида), холецистокинина, секретина, гастрина, желудочного ингибирующего полипептида.
Клинически важным и в какой-то мере опасным является усиление секреции инсулина при длительном воздействии соматотропного гормона, АКТГ и глюкокортикоидов, эстрогенов, прогестинов. При этом возрастает риск истощения β-клеток, уменьшение синтеза инсулина и возникновение инсулинзависимого сахарного диабета. Такое может наблюдаться при использовании указанных гормонов в терапии или при патологиях, связанных с их гиперфункцией.
Нервная регуляция β-клеток поджелудочной железы включает адренергическую и холинергическую регуляцию. Любые стрессы (эмоциональные и/или физические нагрузки, гипоксия, переохлаждение, травмы, ожоги) повышают активность симпатической нервной системы и подавляют секрецию инсулина за счет активации α2-адренорецепторов. С другой стороны, стимуляция β2-адренорецепторов приводит к усилению секреции.
Также выделение инсулина контролируется n.vagus, в свою очередь находящегося под контролем гипоталамуса, чувствительного к концентрации глюкозы крови.
К органам-мишеням инсулина можно отнести все ткани, имеющие к нему рецепторы. Рецепторы инсулина находятся практически на всех клетках, кроме нервных, но в разном количестве. Наибольшая их концентрация наблюдается на мембране гепатоцитов (100-200 тыс на клетку) и адипоцитов (около 50 тыс на клетку), клетка скелетной мышцы имеет около 10 тысяч рецепторов, а эритроциты - только 40 рецепторов на клетку.
Нервные клетки не имеют рецепторов к инсулину, который просто не проникает через гематоэнцефалический барьер.
Рецептор инсулина представляет собой гликопротеин, построенный из двух димеров, каждый из которых состоит из α- и β-субъединиц, (αβ)2. Обе субъединицы кодируются одним геном 19 хромосомы и формируются в результате частичного протеолиза единого предшественника. Период полужизни рецептора составляет 7-12 часов.
При связывании инсулина с рецептором изменяется конформация рецептора и они связываются друг с другом, образуя микроагрегаты.
Связывание инсулина с рецептором инициирует ферментативный каскад реакций фосфорилирования. Первым делом аутофосфорилируются тирозиновые остатки на внутриклеточном домене самого рецептора. Это активирует рецептор и ведет к фосфорилированию остатков серина на особом белке, называемом субстрат инсулинового рецептора (СИР, или чаще IRS от англ. insulin receptor substrate). Таких IRS выделяют четыре типа – IRS‑1, IRS‑2, IRS‑3, IRS‑4. Также к субстратам инсулинового рецептора относят белки Grb-1 и Shc, которые отличаются от IRS аминокислотной последовательностью.
Дальнейшие события разделяются по двум направлениям:
1. Реакции, связанные с активностью ферментов MAP-киназ – в целом управляют активностью хроматина.
2.
Процессы, связанные с
активацией фосфоинозитол-3-киназы –
в основном контролируют метаболические
реакции. Сюда же относятся процессы,
которые регулируют активность глюкозных
транспортеров и
поглощение глюкозы клетками.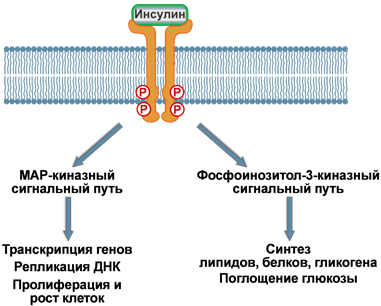
Тем не менее, такое подразделение условно, так как в клетке присутствуют ферменты, чувствительные к активации и того и другого каскадного пути.