

Билеты_бф_экзамен_-_2019
.PDF1.Аутокринная, паракринная и эндокринная регуляция клеточной активности.
По расстоянию от клетки–продуцента гормона до клетки-мишени можно выделить 3 варианта регуляции:
Эндокринная/дистантная – секреция происходит в жидкие среды организма. Клеткимишени при этом могут быть сколько угодно далеко. Пр-р: секреторные клетки эндокринных, гормоны из которых поступают в систему общего кровотока – гипофиз, надпочечники, щитовидная железа, половые железы Паракринная – продуценты БАВ и клетка-мишень расположены рядом. Молекула гормона
достигают мишени путём диффузии в межклеточном веществе. Митогенные стимуляторы эпидермальный фактор роста, фактор роста тромбоцитов, ИЛ-2 (фактор роста Т-клеток) Аутокринная – клетка-продуцент гормона имеет рецепторы к этому же гормону (т.е. клетка продуцент и мишень одновременно). Пр-р: эндотелины, вырабатываемые клетками эндотелия и воздействующие на эти же эндотелиальные клетки, Т-лимфоциты секретирующие интерлейкины
2.Понятие первого и второго посредников. Классификация первых посредников.
Первичные посредники – межклеточные сигнальные молекулы Вторичные (медиаторы, мессенджеры) – внутриклеточные, вырабатываются при
взаимодействии сигнальной молекулы с рецептором мембраны внутри клетки, запускают каскадный механизм, который приводит к изменениям активности клетки
Первичные делятся на:
1.Небольшие липофильные молекулы (стероидные и тиреоидные гормоны) – посредством диффузии проникают через мембрану и связываются с внутриклеточными рецепторами. Имеют гормон-связывающий домен и ДНК-связывающий домен.Итог действия транскрипция определенных генов
2.Липофильные молекулы – производные арахидоновой кислоты – эйкозаноиды (простагландины, лейкотриены, простациклины и тромбоксаны). Образуются практически во всех клетках их синтез инициируется фосфолипазой А2.
Действуют через мембранные рецепторы аутокринно и паракринно. Регулируют сокращения гладкомышечных клеток, болевые и воспалительные реакции, секрецию желудочного сока, агрегацию тромбоцитов
3.Гидрофильные молекулы – гидрофильные гормоны (адреналин), нейротрансмиттеры, цитокины, факторы роста. Вырабатываются специализированными эндокринными железами или клетками, транспортируются к клеткам-мишеням кровотоком. Общий механизм запуск каскадов внутриклеточных сигналов, приводящих к определенным физиологическим эффектам.
3.Компоненты регуляторной системы, связанной с вторичными посредниками. Характеристики вторичных посредников на примере цАМФ
1.Рецептор, специфически узнающий и взаимодействующий с сигнальной молекулой.
2.Трансдуктор сигнала – гетеротримерный ГТФ – связывающий G-белок
3.Эффектор, воспринимающий и усиливающий сигнал
Взаимодействие лиганда с рецептором вызывает запуск каскадного механизма, усиливающего исходный сигнал, что сопровождается образованием вторичного посредника. Внутриклеточный медиатор вызывает активацию соответствующей протеинкиназы, фосфорилирующей определенные белки, что в конечном счете ведет к физиологическому эффекту.
1.Химический агент должен стимулировать активность аденилатциклазы в препаратах чувствительной ткани
2.В ответ на действие хим.агента должна возрастать концентрация цАМФ в ткани
3.Косвенное доказательство участия цАМФ в реакции –потенцирование гормонального эффекта ингибиторами фосфодиэстеразы
4.Воспроизведение гормонального эффекта с помощью цАМФ или его производного
5.Повышение концентрации цАМФ в ткани под влиянием гормона должно предшествовать регистрируемой биологической реакции
4.Классификация рецепторов. Свойства рецептора как компонента сигнальной системы
Рецептор – специфический белок, который связывает сигнальное вещество, что приводит в конечном итоге к развитию клеточной реакции Агонист – активирует, антагонист – дезактивирует
Свойства рецепторов:
Селективность – строгая структурная специфичность. Данный лиганд должен быть либо единственным вещ-вом, связывающимся с рецептором, либо эффективно конкурировать с другими Насыщаемость – число мест связывания с лигандом должно быть конечно
Сродство к лиганду – насыщение рецептора происходит при физиологических концентрациях лиганда Тканевая специфичность – связывание лиганда с рецептором происходит в той ткани, где
наблюдается её биологическое действие Классификация рецепторов:
1.Рецепторы-каналы – связывание с лигандом приводит к изменению конформации рецептора, что позволяет определенным ионам проходить через канал. Результат – изменение мембранного потенциала/внутриклеточного уровня вторичного последника
2.Рецепторы, сопряженные с G-белками. В результате связывания активируется G-белок, который стимулирует/ингибирует ферменты, производящие вторичные посредники
3.Каталитические рецепторы, проявляющие ферментативную активность: гуанилатциклазную, фосфатазную, тирозинкиназную (инсулин, ростовые факторы)
4.Рецепторы не проявляющие ферментативную активность, но сопряженные с тирозинкиназой – связывание приводит к димеризации мономерных рецепторов, кторые активируют тирозинкиназу (рецепторы цитокинов, интерферонов)
Мембранные (внешнеклеточный домен, трансмембранный участок, участок погруженный в цитоплазму специфичен к ассоциированному внутриклеточному белку) и цитоплазматические (представлены глобулярными белками)
Классификация по видам агонистов: адренэргические, холинергические, серотонинэргические
5.ГТФ-связывающие белки: структура, принципы действия, методы изучения
ГТФ-связывающие белки или G-белки являются гетеродимерными. Состоят из альфа, бета и гамма субъединиц.
Взаимодействуют и с рецептором и с эффектором. Альфа-субъединица обладает ГТФ-
азной активностью.
В неактивном состоянии альфа-субъединица связана с ГДФ. Присоединение агониста к рецептору вызывает изменение его конформации, вследствие чего рецептор связывается с G-белком. Образуется комплекс белка и рецептора, и ГДФ меняется на ГТФ.
Белок активируется, отделяется от рецептора и распадается на альфа и бета+гамма субъединицы.
Белки-эффекторы: аденилатциклаза, фосфолипаза С, белки ионных каналов, обменников и др.
Переход G-белка в неактивное состояние происходит благодаря гидролизу ГТФ. Как только ГТФ меняется на ГДФ альфа-субъединица теряет свою активность, отсоединяется от белка-эффектора и ассоциируется с бета+гамма комплексом.
По действию на аденилатциклазу есть два вида белков : активирующие и ингибирующие
Чтобы различить эти белки используют токсины бактерий.
Холерный токсин действует на активирующие белки, а коклюшный на ингибирующие. Они оба содержат фермент АДФ-риболазу.
6.Сигнальная система, опосредованная цАМФ. Аденилатциклаза, фосфодиэстераза, протеинкиназа А
цАМФ – вторичный посредник.
Аденилатциклаза – катализирует превращение АТФ в цАМФ. Является интегральным белком. Активируется при взаимодействии с альфа-субъединицей стимулирующего ГТФсвязывающего белка.
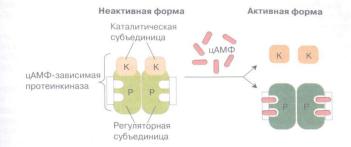
Увеличение в клетке цАМФ активирует цАМФ-зависимую протеинкиназу (протеинкиназу А). Это белок неактивном состоянии состоит из 2 регуляторных и 2 каталитических субъединиц. После присоединения 4 молекул цАМФ к регуляторным субъединицам происходит диссоциация комплекса, и каталитические субъединицы приобретают активность
Инактивирует цАМФ – фосфодиэстераза
7.Сигнальная система, опосредованная цГМФ. Две формы гуанилатциклазы. Протеинкиназы G
Две формы: мембранносвязанная и растворимая.
Первый путь: Рецепторный домен расположен на внешней стороне мембраны, а каталитический на её внутренней стороне. При связывании агониста с рецептором происходит активация мембраносвязанной гуанилатциклазы и нарабобтка цГМФ. Циклический ГМФ связывается с цГМФ-зависимой протеинкиназой, которая, как и протеинкиназа А отнсится к серинтреониновым протеинкиназам.
Состоит из 2 субъединиц, имеет 2 регуляторных центра, с которыми соединяются 2

молекулы цГМФ. При активации субъединицы не разъединяются.
Пример агониста – натрийуретический пептид, пептиды секретируемые яичниками и стимулирующие подвижность сперматозоидов.
Второй путь, связан с растворимой гуанилатциклазой. Ее активация происходит под действием оксида азота. Является гетеродимером и содержит SH-группы, гем цГМФ играет важную роль в регуляции Са2+-гомеостаза в различных типах клеток.
Повышение концентрации цГМФ приводит к понижению концентрации Са2+ в результате
активации Са2+-АТФ-аз
активации Na/Ca-обменника
подавления рецепторзависимого поступления Са2+ в клетку
Эти эффекты опосредованы действием протеинкиназы G на мембранные белки, участвующие в обмене Са2+.
Протеинкиназа G фосфорилирует Na,K-АТФазу эпителиоцитов почечных канальцев, что приводит к торможению насоса. Результат: ослабление реабсорбции ионов натрия, а значит и воды из первичной мочи
Особенности цГМФ-зависимой регуляции
-отсутствуют G-белки
-связывание агониста с рецептором всегда приводит к активации гуанилатциклазы
8.NO в роли регулятора клеточных функций. Образование NO в клетке, 2 формы NOсинтазы
Роль оксида азота:
♦регулятор переноса кислорода эритроцита ♦ участник межнейрональной передачи информации
♦фактор, влияющий на гипотоническую устойчивость эритроцитов ♦участник иммунных реакций ♦эндогенный вазодилятатор ♦регулирует сократимость скелетных мышц ♦ингибирует агрегацию тромбоцитов
NO вырабатывается из L-аргинина под действием NO-синтазы во многих клетках, но существует три категории клеток, в которых проявляются функции этой молекулы: эндотелиальные клетки (расслабление гмк)
нейроны (передача сигнала)
иммунокомпетентные клетки (участие в иммунном ответе)
Эндотелиальная NO-синтаза: нерастворимый фермент с молекулярной массой около 135 кДа. Продуцируется эндотелиальной синтазой. Регулирует тонус кровеносных и лимфатических сосудов, предупреждает тромбообразование
Нейрональная NO-синтаза: растворимый гомодимер с м.м. 150 кДА. Содержит кальмодулин-связывающий центр, осуществляющий Ca-зависимую регуляцию синтеза NO. Содержит атом Fe, входящий в состав гемовой простетической группы.
Контролирует осцилярную активность нейронов, является медиатором ноцицепции, термочувствительности, обоняния, регулирует выход нейромедиаторов, играет центральную роль в процессах обучения и памяти.
Индуцибельная NO-синтаза: кальций-независимая обнаружена в макрофагах, гепатоцитах, фибробластах. Индуцируется в зараженных вирусом клетках и стимулирует образование нестабильных свободных радикалов
Конститутивная форма |
Индуцибельная форма |
В эндотелиальных клетках и нейронах |
В макрофагах |
Активность зависит от Ca2+ |
Активируется цитокинами |
Связана с мембраной |
Цитозольный фермент |
Концентрация образуемого NO невелика |
Концентрация образуемого NO большая |
Функции: расслабление ГМК сосудов, |
Функции: цитостатическое и |
межнейрональная передача |
цитотоксическое действие на атакуемые |
|
макрофагами клетки |
9.Са-зависимая система регуляции. Ионы кальция в роли вторичного посредника. Кальций-связывающий белок кальмодулин и его роль в клетке
Роль ионов кальция в клетках: сокращение мышц, выделение нейромедиаторов,
вторичный посредник
Низкая внутриклеточная концентрация ионов кальция ( 10-7 М) - возможность быстро повысить внутриклеточную концентрацию (до 0,6 – 2 мкМ)
Характеристика Са,как вторичного посредника
1.В ответ на действие БАВ активируются структуры, которые поставляют ионы кальция в клетку
2.В ответ на БАВ увеличивается внутриклеточная концентрация кальция
3.Снижение активности Ca-АТФазы приводит к усилению эффекта ионов Ca
4.Эффект БАВ воспроизводится с помощью Са-ионофора
5.Повышения внутриклеточного кальция предшествует биологическому эффекту Кальмодулин – является кальций-связывающим белком, не проявляющим ферментативной активности.
Впервые обнаружен Ченгом в мозге КРС в качестве Са-зависимого активатора фосфодиэстеразы циклических нуклеотидов.
Впоследствии обнаружен во всех эукариотических клетках.
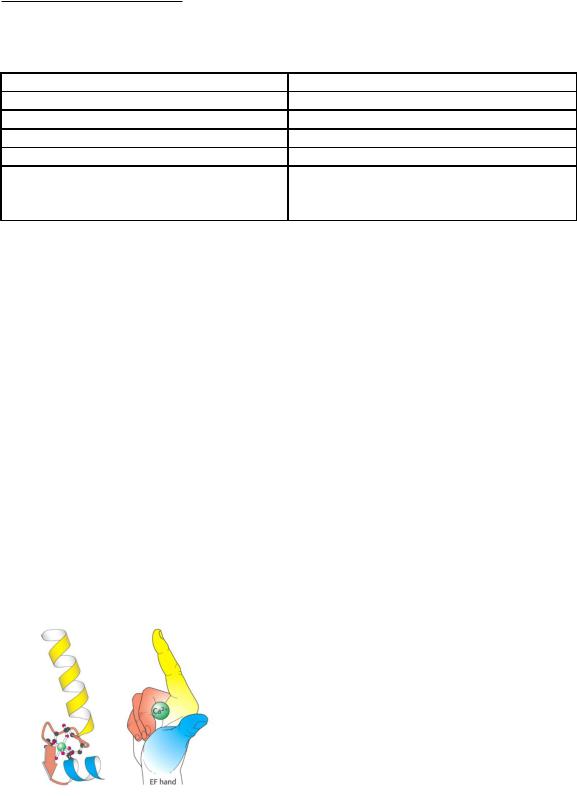
Является «E-F hand» - протеином.
Вструктуре белка было обнаружено 6 -спиралей (A, B,C, D, E, F).
Впетлях полипептидной цепи между участками C и D, E и F находятся функциональные группы с высоким сродством к кальцию.
Спирали E и F расположены как вытянутые указательный и большой палец.
4 Са-связывающих центра, кроме кальмодулина, имеют тропонин С (из скелетных мышц), легкие
цепи миозина, кальцийнейрин Са-кальмодулин-зависимые ферменты: киназа легких цепей миозина (в ГМК), Са-АТФаза, аденилатциклаза, ФДЭ, Са-СаМ-зависимая протеинкиназа
Са-кальмодулин-зависимые процессы: сокращение ГМК, клеточная пролиферация, диссоциация микротрубочек, высвобождение нейромедиаторов Комплекс кальций-кальмодулин: прямое взаимодействие с ферментом-мишенью/через Са-кальмодулин зависимую протеинкиназу
10.Продукты гидролиза фосфоинозитидов в роли вторичных посредников
Инозитол-3-фосфат – гидрофильная молекула, которая диффундирует в цитозоль и взаимодействует с рецепторами ретикулума Диацилглицерол – гидрофобная молекула, которая остается в мембране. Под действием
фосфолипазы А2 дает арахидоновую кислоту, которая является предшественников эйказаноидов
11.Две формы фосфолипазы С (β,γ). Протеинкиназа С и ее роль в регуляции клеточных функций.
Фосфолипаза С-бета активируется через рецепторы, связанные с G-белками (Gq белок) α1-адренорецепторы, мускариновые рецепторы Фосфолипаза С-гамма активируется благодаря фосфолированию её тирозинкиназой: факторы роста
Протеинкиназа С участвует в таких процессах, как: секреция, сокращения ГМК, агрегация тромбоцитов, фосфорилирование рецепторов Состоит из 2 доменов – регуляторного и каталитического.
Каталитический домен: центр, связывающий АТФ и белок-субстрат Регуляторный домен: мотив цинковые пальцы и участок с высоким сродством к Са Неактивная протеинкиназа С – цитозольная, при активации становится мембранносвязанной
12.Рецепторные тирозинкиназы. Ras-белок
Агонисты для рецепторных тирозинкиназ: фактор роста/митогены и инсулин Состоит из внеклеточного N-концевого участка, связывающего агонисты, трансмембранного участка, внутриклеточного тирозинкиназного домена
Классификация тирозинкиназных рецепторов: Группа рецепторов с участками, богатыми цистеином
1.Семейство рецепторов эпидермального фактора роста
2.Семейство рецепторов инсулина и инсулиноподобного фактора роста
3.Семейство рецепторов фактора роста гепатоцитов
Семейство с иммуноглобулинподобными доменами во внеклеточной области и дополнительным регуляторным участком в тирозинкиназном домене
1.Рецепторы фактора роста тромбоцитов содержат 5 таких доменов
2.Рецепторы фактора роста фибробластов содержат 3 домена
Механизм функционирования рецепторной тирозинкиназы (РТК) Димеризация рецепторов Трансавтофосфорилирование тирозиновых остатков
Формирование участков для связывания белков-субстратов, содержащих SH2 и SH3 домены Фосфорилирование связывающихся с РТК белков

Ключевой компонент сигнального пути, связанного с рецепторной тирозинкиназой – Ras- белок
Ras-белок – мономерная ГТФаза
В активном состоянии Ras-белок контактирует с ГТФ, в неактивном с ГДФ.
Активация и инактивация осуществляется двумя специфическими белками GEF и GAP После активации в процесс включается несколько внутриклеточных киназ (киназный каскад)
Эффект – деление клеток
13.Монооксид углерода, сероводород как регуляторные молекулы: образование в клетках, физиологическое значение
Субстрат для образования СО – молекула гема, являющаяся небелковым компонентом гем-протеинов (гемоглобин, миоглобин, каталаза, пероксидаза, цитохром). Реакция катализируется гем-оксигеназой и протекает по уравнению:
гем --- (NADPH) --- > биливердин + СО + Fe2+
Функции СО:
-нейтротрансмиттер
-влияние на тонус гладкой мускулатуры сосудов
-противосвертывающий эффект
-сигнальная роль в иммунной системе
Сероводород: образуется из L-цистеина цитозольными пиродоксаль-5-фосфат (витамин В6) – зависимыми ферментами – цистатионин -синтазой (CBS) и цистатионин -лиазой (CSE);
Неферментативный – синтез H2S при окислении глюкозы в присутствии элементарной серы Эффекты сероводорода – эндогенный вазодилятатор, ингибирует агрегацию тромбоцитов, регуляция нейрональной активности, провоспалительный агент, поскольку усиливает выработку провоспалительных цитокинов
14.Электропроводность биообъектов для постоянного тока. Токи смещения и токи проводимости. Видимое отклонение от закона Ома
Биологические ткани – композиционные среды, т.к. имеют элементы со свойствами как проводников, так и диэлектриков Проводники – вещества, в которых СВОБОДНЫЕ заряды перемещаются по всему объёму.
Свободные заряды – заряженные частицы одного знака, способные перемещаться под действием электрического поля Диэлектрики – вещества, содержащие только СВЯЗАННЫЕ заряды
Связанные заряды – разноимённые заряды, входящие в состав атомов и молекул, которые НЕ МОГУТ перемещаться под действием поля независимо друг от друга.
Ток проводимости - перемещение СВОБОДНЫХ зарядов под действием поля Ток смещения – перемещение СВЯЗАННЫХ зарядов
Электропроводность – свойство живого тела пропускать электрический ток под воздействием электрического поля. Обусловлена наличием свободных зарядов в ткани. Существенно зависит от содержания в ткани воды Измеряется в СИМЕНСАХ 1 См = 1/Ом = = А/В = кг-1 м-2 с3 А2

Отклонение от закона Ома
При пропускании постоянного электрического тока через биологические объекты его величина с течением времени уменьшается, достигая уровень намного ниже исходного. Причина: вектор напряженности Еп элетрического поля, создаваемого связанными зарядами на поверхности диэлектрика, направлен внутри диэлектрика противоположно вектору напряженности Ео внешнего поля, вызывающего поляризацию
I измеряется в АМПЕРах, R измеряется в Омах, P(t) – ЭДС поляризации зависит от времени
Электродвижущей силой источника (сокращенно ЭДС) называется скалярная физическая величина – количественная мера способности источника создавать на полюсах разность потенциалов. Она равна работе сторонних сил по перемещению заряженной частицы с положительным единичным зарядом от одного полюса источника к другому, т.е.
В СИ ЭДС измеряется в вольтах (В), т.е. в тех же единицах, что и напряжение
15.Явление поляризации, её виды Поляризация – процесс перемещения связанных зарядов под действием внешнего
электрического поля и создание вследствие этого ЭДС, направленной против внешнего поля. Наиболее выражено при измерении сопротивления на постоянном токе Виды:
Электронная – упругое смещение электронных орбит относительно ядер в атомах и молекулах под действием внешнего электрического поля (время релаксации 10 -16 – 10 -14 с) Ионная – упругое смещение противоположно заряженных ионов в узлах кристаллической решетки. Присутствует в кристаллических веществах. Ионная и электронная поляризация происходит без потерь энергии (10 -16 – 10 -12 с)
Дипольная – полярные молекулы имеют несимметричное строение. Центры тяжести разноименных зарядов у них не совпадают, и поэтому в отсутствие внешнего электрического поля эти молекулы представляют собой диполи (10 -13 – 10 -7 с) Макроструктурная – возникает под действием электрического поля, вследствие неоднородности электрических свойств вещества (10 -8 – 10 -3 с)
Электролитическая поляризация – связана с поляризацией электродов, опущенных в раствор электролита при пропускании через них тока (до нескольких секунд)
16.Электропроводность биобъектов для переменного тока. Суммарное сопротивление, его
определение при последовательном и параллельном соединениях омического и реактивного сопротивлений. Дисперсия импеданса. Коэффициент Тарусова.
- Сопротивление биологических структур переменному току НИЖЕ, чем постоянному току - Сопротивление НЕ ЗАВИСИТ ОТ ВЕЛИЧИНЫ

тока, если эта величина не превышает физиологическую норму
-На ФИКСИРОВАННОЙ ЧАСТОТЕ тока сопротивление биологического объекта ПОСТОЯННО, если не изменяется его физиологическое состояние
-Сопротивление существенно ИЗМЕНЯЕТСЯ при изменении ФИЗИОЛОГИЧЕСКОГО состояния биообъекта, например, при гибели (отмираниии) сопротивление падает на несколько порядков
-ЭЛЕКТРОПРОВОДНОСТЬ биологических объектов с УВЕЛИЧЕНИЕМ частоты УВЕЛИЧИВАЕТСЯ
-ДИСПЕРСИЯ ЭЛЕКТРОПРОВОДНОСТИ, как и способность к поляризации, присущи только живым клеткам
Дисперсия импеданса – зависимость суммарного сопротивления от частоты переменного тока
Коэффицинт Тарусова – дисперсия электропроводности характерна только для живых объектов. По мере отмирания тканей резко увеличивается низкочастотная (при той же высокочастотной) компонента. Для оценки жизнеспособности Б.Н. Тарусовым был предложен коэффициент
К = R*10^4/R*10^6
числитель – сопротивление при частоте тока в 10^4 Гц знаменатель – сопротивление при частоте тока в 10^6 Гц При отмирании тканей К стремится к 1
Определение коэффициента Тарусова позволяет объективно оценить способы консервации, условий и сроков хранения различных тканей, предназначенных для трансплантации.
17.Раздражимость и возбудимость. Порог возбуждения. Крива сила – длительность. Реобаза, хронаксия.
Раздражимость – способность живого организма реагировать на внешнее воздействие окружающей среды изменением своих физико-химических и физиологических свойств. Возбуждение – сложный биологический процесс, который характеризуется специфическим изменением обмена веществ, временной деполяризацией мембраны клеток и проявляющейся специализированной реакцией ткани.
Порог раздражения – мера возбудимости, минимальная сила раздражителя, вызывающая ответ.
Существует определенное соотношение между временем действия раздражителя и его амплитудой. Эта зависимость получил название кривой «сила-длительность»
На этой кривой видно, что уменьшение значения тока ниже определенной критической величины не приводит к возбуждению ткани независимо от продолжительности времени, в течение которого действует этот раздражитель

Реобаза – минимальная величина тока, вызывающая возбуждение. Определяется разностью между критическим потенциалом и мембранным потенциалом покоя.
Полезное время – в течении которого должен действовать раздражитель пороговой силы с тем, чтобы вызвать возбуждение
Порог времени определяется емкостью и резистивной характеристикой клеточной мембраны. Т.е. постоянной времени T=R*C
Хронаксия – время, в течении которого должен действовать раздражитель удвоенной реобазы
18.Потенциал покоя и механизм его формирования. Уравнение Нернста, уравнение Гольдмана
По предположению Бернштейна поверхностная мембрана возбудимой клетки в покое обладает избирательной проницаемостью: ионы К проходят через неё гораздо легче, чем ионы Na и Сl. Т.к. концентрация К в клетке выше, чем во внеклеточной среде, диффузия этих ионов через мембрану создаёт на ней РАЗНОСТЬ ПОТЕНЦИАЛОВ – потенциал покоя (ПП) причём внутренняя сторона мембраны оказывается заряженной отрицательно, а внешняя – положительно.
Существуют такие понятия, как:
Диффузионный потенциал: подвижность ионов хлора (v) выше, чем ионов натрия (u)
Мембранный потенциал – мембрана непроницаема для ионов хлора
e – внешняя концентрация ионов, I – внутренняя
концентрация ионов
Электрохимическое равновесие на полупроницаемой мембране возникает благодаря диффузионному давлению в равновесии с противоположной электростатической силой,