

2.6 Способ записи генетической информации в молекуле днк. Биологический код и его свойства.
Генетический код (он же биологический)- способ записи наследственной информации в молекулах нуклеиновых кислот в виде последовательности образующих эти кислоты нуклеотидов.
Генети́ческий код — свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
В ДНК используется четыре азотистых основания — аденин (А), гуанин (G), цитозин (С), тимин (T). Эти буквы составляют алфавит генетического кода. В РНК используются те же нуклеотиды, за исключением тимина, который заменён похожим нуклеотидом — урацилом. В молекулах ДНК и РНК нуклеотиды выстраиваются в цепочки и, таким образом, получаются последовательности генетических букв.
![]()
Генетический код
Белки практически всех живых организмов построены из аминокислот всего 20 видов. Эти аминокислоты называют каноническими. Каждый белок представляет собой цепочку или несколько цепочек аминокислот, соединённых в строго определённой последовательности. Эта последовательность определяет строение белка, а следовательно все его биологические свойства.
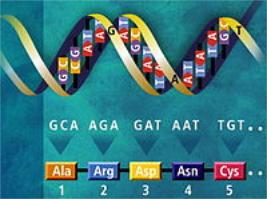
Реализация генетической информации в живых клетках (то есть синтез белка, кодируемого геном) осуществляется при помощи двух матричных процессов: транскрипции (то есть синтеза мРНК на матрице ДНК) и трансляции генетического кода в аминокислотную последовательность (синтез полипептидной цепи на мРНК). Для кодирования 20 аминокислот, а также сигнала «стоп», означающего конец белковой последовательности, достаточно трёх последовательных нуклеотидов. Набор из трёх нуклеотидов называется триплетом. Принятые сокращения, соответствующие аминокислотам и кодонам, изображены на рисунке.
Свойства
1. Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
2. Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно.
3. Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).
4. Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин)[1]
5. Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов.
6. Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии).
7. Помехоустойчивость — мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.
2.7 Механизмы сохранения нуклеотидной последовательности днк.
Устойчивость к внешнему воздействию
ДНК химически инертные вещество. Роль передачи наследственной информации может выполнять и РНК (некоторые вирусы). Считают, что выбор в пользу ДНК обусловлен её более низкой (по сравнению с РНК) реакционной способностью.
Механизм коррекции ошибок
Точность копирования нуклеотидных последовательностей материнской ДНК в процессе ее репликации.
Сам процесс репликации по пунктам:
ДНК-геликаза расплетает двойную спираль ДНК, разделяя ее полинуклеотидные цепи;
дестабилизирующие белки выпрямляют участок цепи ДНК;
ДНК-топоизомераза разрывает фосфодиэфирную связь в одной из полинуглеотидных цепей ДНК, снимая напряжение, вызываемое расплетенисм спирали и расхождением цепей в репликационной вилке;
РНК-праймаза синтезирует РНК-затравки для дочерней цепи и для каждого фрагмента Оказаки;
ДНК-полимераза осуществляет непрерывный синтез лидирующей цепи и синтез фрагментов Оказаки отстающей цепи;
ДНК-лигаза сшивает фрагменты Оказаки после удаления РНК-затравки.
Рассмотренный выше механизм репликации отличается чрезвычайно высокой точностью воспроизведения структуры ДНК. При удвоении ДНК ошибки возникают в среднем с частотой 1·10-6 комплементарных пар оснований.
В поддержании высокой точности репликации важная роль принадлежит прежде всего ферменту ДНК-полимеразе. Этот фермент осуществляет отбор необходимых нуклеотидов из числа имеющихся в ядерном соке нуклеозидтрифосфатов (АТФ, ТТФ, ГТФ, ЦТФ), точное присоединение их к матричной цепи ДНК и включение в растущую дочернюю цепь (см. рис. 3.10). Частота включения неправильных нуклеотидов на этой стадии составляет 1·10-5 пар оснований.
Эксцизионная репарация («Вырезание»). Осуществляется до следующей репликации, поэтому называют так же дорепликативной.
Причина ошибки |
|
Описание |
Ферменты |
Коррекция |
Возникновение измененных форм азотистых оснований |
Ц`-A |
Ц быстро переходит в исходную форму и связь разрушается, появляется неспаренный 3'-ОН-конец |
ДНК-полимераза, Эндонуклеаза
|
Отщепление ошибочно включенного в цепь ДНК нуклеотида |
Нарушение концентрации нуклеозидтрифосфатов |
А > Т Г > Ц И наоб. |
выпадению пар оснований / замена одних пар другими ??? |
(не указаны) |
|
Потеря пуриновых оснований |
-Г -Ц |
Выпадения пар оснований |
Эндонуклеаза |
Разрывает фосфоэфирную связь в месте повреждения цепи. |
Экзонуклеаза |
Удаляет измененный участок с несколькими примыкающими к нему нуклеотидами |
|||
Дезаминирование Цитозина, Алкилирование и др |
Ц->У |
Замена одних пар другими |
ДНК-гликозилаза |
Модифицированные основания удаляются. Возникшие пробелы заполняются по принципу комплементарности. * |
Ультрафиолетовое излучение |
Т-Т |
Замена одних пар другими |
(не указаны) |
Удаление участка, несущего димер, и восстановлением нормальной последовательности нуклеотидов |
* Если восстановление нормальной структуры не осуществляется, например в случае дезаминирования азотистых оснований, происходит замена одних пар комплементарных оснований другими — пара Ц—Г может заменяться парой Т—А и т.п.
Когда дорепликативная репарация не устраняет ошибки происходит фиксация этого изменения (в обеих цепях ДНК). Это приводит к замене одной пары комплементарных нуклеотидов на другую либо к появлению разрывов (брешей) во вновь синтезированной цепи против измененных участков.
Пострепликативная репарация осуществляется рекомбинацией (обмена фрагментами) между двумя вновь образованными двойными спиралями ДНК. Возникшие тиминовые димеры (Т—Т), когда они не устраняются самопроизвольно под действием видимого света (световая репарация) или в ходе дорепликативной эксцизионной репарации, образуют ковалентные связи которые не дают им связаться с Аденином.
В результате во вновь синтезируемой цепи ДНК появляются разрывы (бреши), узнаваемые ферментами репарации. Восстановление целостности новой полинуклеотидной цепи одной из дочерних ДНК осуществляется благодаря рекомбинации с соответствующей ей нормальной материнской цепью другой дочерней ДНК. Образовавшийся в материнской цепи пробел заполняется затем путем синтеза на комплементарной ей полинуклеотидной цепи (рис. 3.16).
Если после до- и пострепликативной репарации остаётся много повреждений включается система индуцируемых (побуждаемых) ферментов репарации (SOS-система). Эти ферменты заполняют бреши, восстанавливая целостность синтезируемых полинуклеотидных цепей без точного соблюдения принципа комплементарности. Вот почему иногда сами процессы репарации могут служить источником стойких изменений в структуре ДНК (мутаций).
Если в клетке, несмотря на осуществляемую репарацию, количество повреждений структуры ДНК остается высоким, в ней блокируются процессы репликации ДНК. Такая клетка не делится, а значит, не передает возникших изменений потомству.
Вызываемая повреждениями ДНК остановка клеточного цикла в сочетании с невозможностью молекулярной репарации измененного наследственного материала может с участием белка, синтез которого контролируется геном р53, приводить к активации процесса самоликвидации (апотпоз) дефектной клетки с целью устранения ее из организма.