

Мобільні елементи в геномах: типи та молекулярні механізми переміщення.
Інтерсперсні (мобільні) елементи, здатні до переміщення та розмноження в межах геному (~44 % у геномі людини). Значна частина таких послідовностей є результатом колишньої активності мобільних елементів (таких, що втратили здатність до переміщення), але деякі зберігають свою активність досі. Основні типи мобільних елементів у геномі людини:
• ДНК-транспозони (3 %) переміщення здійснюється шляхом вирізання ділянки ДНК із наступним вбудовуванням її в інше місце.
• LTR-ретропозони (8 %) - як і для інших двох типів мобільних елементів, переміщення відбувається через проміжну молекулу РНК: на елементі послідовності (який містить довгі кінцеві
повтори - Long Terminal Repeats - і кілька генів, зокрема ген зворотної транскриптази) здійснюється транскрипція; молекула РНК прямує до цитоплазми, де відбувається зворотна
транскрипція - синтез ДНК на РНК-матриці за допомогою зворотної транскриптази (РНК-залежна ДНК-полімераза). ДНК повертається до ядра, де вбудовується в геном.
• Мобільні елементи LINE (Long INterspersed Elements, 20 %) містять кілька генів, включаючи ген зворотної транскриптази. На відміну від LTR-ретропозонів, зворотна транскрипція здійснюється в ядрі. У геномі людини присутні три родини таких елементів, одна з них є активною і слугує основним джерелом зворотної транскриптази у клітині.
• Мобільні елементи SINE (Short INterspersed Elements, 13 %) - короткі (100-400 пар основ) беззмістовні елементи, які використовують для переміщення систему LINE. До цього класу належить і Alu-повтор (від назви відповідної рестриктази, яка здатна специфічно гідролізувати цей елемент послідовності).
Мобільні елементи розподілені в геномі нерівномірно: є довгі ділянки, що на 90 % складаються з цих елементів, і такі, де інтерсперсні елементи відсутні. Загалом спостерігається негативна кореляція між щільністю генів і мобільних елементів. Винятком із цієї закономірності є позитивна кореляція між щільністю генів і елементів типу SINE.
Переміщення (транспозиції) мобільних (інтерсперсних) елементів є ще одним особливим типом рекомбінації ДНК, яка відбувається в усіх організмів. Мобільні елементи можна поділити на два основні класи: ДНК-транспозони, що переміщуються за принципом "cut and paste", і елементи, переміщення яких відбувається шляхом синтезу РНК із наступним синтезом ДНК на РНК-матриці.
Основними типами мобільних елементів, які належать до другого класу, є: LTR-ретропозони з довгими кінцевими повторами (Long Terminal Repeats); елементи LINE (Long INterspersed Elements); елементи SINE (Short INterspersed Elements). Переміщення мобільних елементів є досить рідкою подією - у бактерій відбувається одна транспозиція на 105 -106 клітин. Проте активність мобільних елементів має важливе біологічне значення. Вбудовування мобільних елементів у кодуючі частини генів очевидно приводить до мутацій, у регуляторні - до зміни транскрипційної активності. У процесі свого переміщення мобільний елемент може захопити той чи інший регуляторний елемент послідовності та вбудувати його в інший промотор. Переміщення генетичного матеріалу разом із мобільними елементами розглядається сьогодні як один із найважливіших еволюційних факторів.
ДНК-транспозони
Мобільні елементи цього типу часто містять один або два гени, що кодують один або два білки під спільною назвою транспозаза. Транспозаза здійснює каталіз хімічних реакцій, що забезпечують транспозицію елемента - його вирізання з донорного сайта та вбудовування в сайт-мішень. Крім генів транспозази мобільний елемент може містити інші гени, або навпаки - гени транспозази можуть бути пошкодженими. В останньому випадку транспозиція даного елемента відбувається з використанням транспозази, закодованої іншим ДНК-транспозоном. Кодуюча частина транспозона фланкована невеликими повторами, котрі розпізнає транспозаза, вирізаючи фрагмент ДНК з тупими кінцями (розрізи у двох полінуклеотидних ланцюгах розташовані точно один напроти одного). Сайт-мішень - невелика специфічна послідовність ДНК, яка теж упізнається транспозазою і теж розрізається - але так, що в місці розрізу залишаються одно ланцюгові 5'-кінцеві вирости. Далі транспозаза каталізує ковалентне приєднання 3'-кінців транспозона до цих виростів, прогалини заповнюються шляхом репатаривного синтезу. Процес транспозиції (який, по суті, є одним із варіантів сайт-специфічної рекомбінації) залишає дволанцюговий розріз у місці, де знаходився транспозон. У випадку незалежної від реплікації транспозиції (нереплікативна транспозиція), цей розріз піддається репарації за механізмом негомологічного з'єднання кінців, тобто транспозон просто «стрибає» в інше місце. Але досить часто транспозиція відбувається під час реплікації (реплікативна транспозиція) - тоді розріз репарується за рекомбінаційним механізмом: сестринська молекула ДНК використовується як матриця, і ділянка, що містила транспозон, відновлюється - транспозон «розмножується».
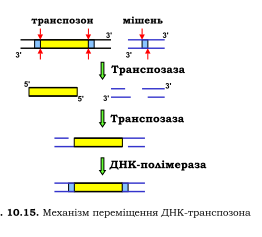
LTR-ретропозони
Кодуюча ділянка LTR-ретропозона (5-8 тис. пар основ) містить кілька генів: зворотної транскриптази (РНК-залежна ДНК-полімераза), інтегрази (аналог транспозази), РНКази Н. На кінцях розташовані довгі кінцеві повтори (200-600 пар основ), які й надали назву цим мобільним елементам.

Процес переміщення копії LTR-ретропозона нагадує життєвий цикл ретровірусів. Першим етапом є транскрипція ретропозона: синтез мРНК РНК-полімеразою ІІ. Ця мРНК транспортується до цитоплазми, де піддається трансляції. Зворотна транскриптаза, яка є продуктом цієї трансляції, здійснює синтез ДНК: мРНК використовується як матриця, 3'-кінець молекули тРНК - як праймер. Комплекс синтезованої ДНК-копії ретропозона з інтегразою повертається до ядра, де здійснюється інсерція цієї ДНК у геном.
Мобільні елементи LINE та SINE
Мобільні елементи LINE довжиною приблизно 6 тис. пар основ містять принаймні два гени -зворотної транскриптази та ендонуклеази - і невеликі специфічні послідовності на кінцях. Процес переміщення копії елемента починається так само, як і у випадку ретропозонів - із транскрипції РНК-полімеразою ІІ. Після трансляції в цитоплазмі синтезовані білки зв'язуються з мРНК, цей комплекс повертається до ядра, де й відбувається зворотна транскрипція: ендонуклеаза
здійснює одноланцюговий розріз у геномній ДНК, 3'-кінець у місці розрізу використовується як праймер. Після синтезу другого ланцюга ДНК нова копія елемента остаточно вбудовується в геном.
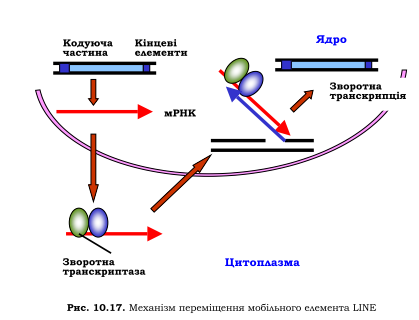
Синтез мРНК при транскрипції елемента LINE, як і для більшості інших еукаріотичних мРНК, закінчується на polyA-сигналі. Цей polyA-сигнал є слабким, що дозволяє елементу вбудовуватись в інтрони звичайних генів без особливих перешкод для експресії цих генів: система процесингу часто не помічає слабкий polyA-сигнал. Елементи LINE є, відповідно, найпоширенішими мобільними елементами в геномі вищих еукаріотів (~20 % геному людини представлено послідовностями LINE) і основним джерелом зворотної транскриптази у клітинах. З часом накопичуються мутації й окремі елементи LINE стають неактивними (тобто вже не кодують зворотну транскриптазу), але переміщуються, використовуючи білки, що кодуються активними елементами. Найчастіше в геномі людини зустрічається елемент типу LINE, котрий позначається як L1 (кілька сотень тисяч копій). Іноді мобільні елементи є не просто ділянками «егоїстичної ДНК», що автономно розмножуються в геномі, а виконують певні конкретні біологічні функції. Наприклад, у дрозофіли відсутня теломеразна система й елемент LINE певного типу виконує функцію подовження кінців хромосом після реплікації: зворотна транскриптаза виступає як теломераза, мРНК мобільного елемента - як теломеразна матрична РНК. Цікаво, що послідовності ДНК гена теломерази та елементів LINE характеризуються високою гомологією: цілком можливо, що теломеразна система походить від мобільних елементів LINE.
Мобільні елементи SINE - короткі (100-300 пар основ) елементи послідовності, які не кодують білків і містять лише промотор для РНК-полімерази ІІІ. Процес їхнього переміщення є таким: елементи SINE «паразитують» на системі LINE, використовуючи її ферменти. На відміну від усіх інших мобільних елементів, елементи SINE сконцентровані в ділянках геному, збагачених на гени. Імовірною причиною цієї закономірності є те, що РНК-продукт SINE має спорідненість до певної протеїнкінази, яка гальмує процес трансляції. Відповідно, зв'язування РНК із протеїнкіназою блокує останню, що приводить до загального підвищення рівня білкового синтезу. Таким чином, підвищення рівня транскрипції SINE разом з активацією інших генів стимулює трансляцію.