

2.1. Физиология нейрона.
Основными элементами нейронной системы являются нервные клетки. Подтверждение клеточной теории строения нервной системы было получено с помощью электронной микроскопии, показавшей, что мембрана нервной клетки напоминает основную мембрану других клеток. Она представляется сплошной на всем протяжении поверхности нервной клетки и отделяет ее от других клеток. Каждая нервная клетка является анатомической, генетической и метаболической единицей так же, как и клетки других тканей организма. Понятие, что одиночная нервная клетка служит, основной функциональной единицей, сменилось представлением о том, что такой функциональной единицей является ансамбль тесно связанных друг с другом нейронов. Нервная система состоит из популяций таких единиц, которые организованы в функциональные объединения разной степени сложности. В нервной системе человека содержится около 100 млрд нервных клеток. Поскольку каждая нервная клетка функционально связана с тысячами других нейронов, то количество возможных вариантов таких связей близко к бесконечности. Нервную клетку следует рассматривать как один из уровней организации нерв-: ной системы, связующих молекулярный, синаптичее-кие, субклеточные уровни с надклеточными уровнями локальных нейронных сетей, нервных центров и функциональных систем мозга, организующих поведение.
Нервные клетки выполняют ряд общих неспецифических функций, направленных на поддержание собственных процессов организации. Это обмен веществами с окружающей средой, образование и расходование энергии, синтез белков и др. Кроме того, нервные клетки выполняют свойственные только им специфические функции по восприятию, переработке и хранению информации. Нейроны способны воспринимать информацию, перерабатывать (кодировать) ее, быстро передавать информацию по конкретным путям, организовывать взаимодействие с другими нервными клетками, хранить информацию и генерировать ее. Для выполнения этих функций нейроны имеют полярную организацию с разделением входов и выходов и содержат ряд структурно-функциональных частей.
Тело нейрона, которое связано с отростками, является центральной частью нейрона и обеспечивает питанием остальные части клетки. Тело покрыто слоистой мембраной, которая представляет собой два слоя липидов с противоположной ориентацией, образующих матрикс, в который заключены белки. Часть мембранных белков является гликопротеинами с полиса-харидными цепочками, выступающими над наружной поверхностью мембраны. Они вместе с углеводами образуют гликокаликс — тонкий слой на поверхности клеточной мембраны, который заполняет межклеточные щели и способствует созданию связей между нейронами, распознаванию клеток, регуляции диффузии через мембрану, обмену с внешней средой. Тело нейрона имеет ядро или ядра, содержащие генетический материал (рис. 2.1),
Ядро регулирует синтез белков во всей клетке и контролирует дифференцирование молодых нервных клеток. При усилении активности нейрона увеличивается площадь ядра и активизируются ядерно-
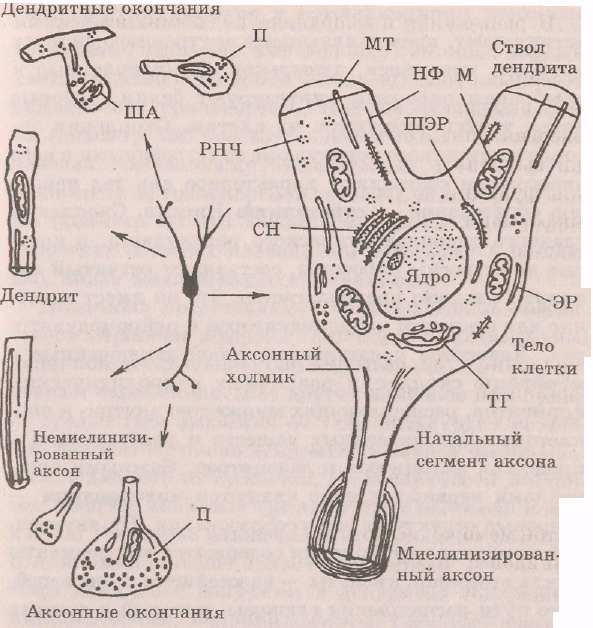
Рис 2.1. Схемы структурно-функциональных частей нейрона. Нейрон, показанный в центре, окружен схемами, иллюстрирующими ультраструктуру его частей: ЭР — эндоплазмати-ческий ретикулюм; ШЭР — шероховатый эндоплазматичес-кий ретикулюм; ТГ ~ тельце Гольджи; СН — субстанция Ниссля; МТ — микротрубочка; НФ — пейрофиламент; М — митохондрия; РНЧ -- рибонуклеопротеиновые частицы; ША — шипиковый аппарат; П — пузырьки (Шеперд, 1987)
плазменные отношения. В цитоплазме тела нейрона содержится большое количество рибосом. Одни рибосомы располагаются свободно в цитоплазме по одной или образуют скопления -- «розетки», где синтезируются белки, которые остаются в клетке. Другие
рибосомы прикрепляются к эидонлазматическому
ретикулюму, представляющему внутреннюю систему мембран, канальцев, пузырьков. Прикрепленные к мембранам рибосомы синтезируют белки, которые потом транспортируются из клетки. Скопления эн-доплазматического ретикулюма со встроенными в него рибосомами составляют характерное для тел нейронов образование — субстанцию Ниссля. Скопления гладкого эндонлазматического ретикулюма, в которые не встроены рибосомы, составляют сетчатый аи-парат Голъджи; предполагается, что он имеет значение для секреции неиромедиаторов и нейромодулято-ров. Лизосомы представляют собой заключенные в мембраны скопления различных гидролитических ферментов, расщепляющих множество внутри- и вне-клеточ ноле каля зова иных веществ и участвующих в процессах фагоцитоза и экзоцитоза. Важными орга-неллами нервных клеток являются митохондрии — основные структуры энергообразования. На внутренней мембране митохондрии содержатся все ферменты цикла лимонной кислоты — важнейшего звена аэробного пути расщепления глюкозы, который в десятки раз эффективней анаэробного пути. Ферменты цепи переноса электронов создают энергию, которая идет на образование АТФ и АДФ. Важной особенностью анергетического обмена нервных клеток является отсутствие собственных углеводов в форме гликогена. Нейроны позвоночных используют глюкозу, беспозвоночных — трегалозу. Высокий уровень энерготрат нервных клеток и отсутствие собственных запасов углеводов делают их особо чувствительными к нарушению поступления крови, в которой содержится глюкоза и кислород, необходимые для аэробного энергообразования на митохондриях. В нервных клетках содержатся также микротрубочкк, нейрофиламенты и
микрофиламенты, различающиеся диаметром. Мжк-ротрубочки (диаметр 300 нм) идут от тела нервной клетки в аксон и дендриты и представляют собой внутриклеточную транспортную систему. Нейрофиламенты (диаметр 100 нм) встречаются только в нервных клетках, особенно в крупных аксонах» и тоже составляют часть ее транспортной системы. Микрофиламен-ты (диаметр 50 нм) хорошо выражены в растущих отростках нервных клеток, они участвуют в некоторых видах межнейронных соединений.
Дендриты представляют собой древовидно»ветвящиеся отростки нейрона, его главное рецептивное поле, обеспечивающее сбор информации, которая поступает через синапсы от других нейронов или прямо из среды. При удалении от тела происходит ветвление дендритов: число дендритных ветвей увеличивается, а диаметр их сужается. На поверхности дендритов многих нейронов (пирамидные нейроны коры„ клетки Пуркинье мозжечка и др.) имеются шипики. Шипиковый аппарат является составной частью системы канальцев дендрита: в дендритах содержатся микротрубочки, нейрофиламенты, сетчатый аппарат Гольджи и рибосомы. Функциональное созревание и начало активной деятельности нервных клеток совпадает с появлением шипиков; продолжи тельное прекращение поступления информации к нейрону ведет к рассасыванию шшшков. Наличие шипиков увеличивает воспринимающую поверхность дендритов; так, площадь дендритов клеток Пуркжнье мозжечка около 250 000 мкм2. Мембрана дендритов по своим свойствам отличается от мембраны других участков нервной клетки и не способна к быстрому и надежному проведению возбуждения.
Аксон представляет собой одиночный,, обычно длинный выходной отросток нейрона, служащий для быс-
трого проведения возбуждения. (В структуру аксона входят начальный сегмент, аксональное волокно и телодендрий.) Аксональное волокно отличается постоянством диаметра по всей длине. В конце он может ветвиться на большое (до 1000) количество веточек. Аксоплазма содержит множество микротрубочек и нейрофиламентов, с помощью которых осуществляется аксональныи транспорт химических веществ от тела к окончаниям (ортоградный) и от окончаний к телу нейрона (ретроградный). Существует быстрый аксональныи транспорт со скоростью сотен миллиметров в сутки и медленный транспорт со скоростью несколько миллиметров в сутки. По аксону транспортируются вещества, необходимые для синаптической передачи, пептиды, продукты нейросекреции. В зависимости от скорости проведения возбуждения различают несколько типов аксонов, отличающихся диаметром, наличием или отсутствием миелиновой оболочки и другими характеристиками (табл. 2.1).
Таблица 2.1 Характеристика разных типов аксонов
Тип аксона |
Скорость, м/с |
Диаметр, мкм |
Наличие миелиновой оболочки |
А а |
420-70 |
22-12 |
Есть |
Ар |
70-40 |
12-8 |
» |
Ау |
40-15 |
8-4 |
» |
А8 |
15-6 |
4-1 |
» |
В |
18-3 |
3-1 |
» |
С |
3-0,5 |
2-0,5 |
Нет |
Начальный сегмент аксона нейронов является тригерной зоной -- местом первоначальной генерации возбуждения. Этот участок нервной клетки начинается от аксонного холмика и, воронкообразно
сужаясь, переходит в начальный участок аксона, не покрытый миелиновой оболочкой. Поскольку этот участок мембраны нейрона является наиболее возбудимым, то здесь обычно первоначально и возникает возбуждение, которое затем распространяется по аксону и телу нейрона. Таких запускающих возбуждение участков может быть несколько. Начальный сегмент аксона имеет важное значение для интегратив-ной деятельности нервной клетки. Телодендрий представляет собой часть нервной клетки, которая осуществляет соединение с другими нейронами путем синаптических контактов. Это конечные разветвления — терминали аксона, которые не покрыты миелиновой оболочкой и заканчиваются утолщениями различной формы (булавы, кольца/пуговки, чаши и др.), которые входят составной частью в синапс. В утолщениях локализовано значительное количество пузырьков, расположенных свободно или встроенных в пресинаптические мембраны. Поскольку терминали аксона очень тонкие и не покрыты миелином, то скорость возбуждения в них значительно меньше, чем в аксонах.
Взаимодействие частей нервных клеток обеспечивает реализацию их функций с помощью химических и электрических процессов. Химические процессы в нервных клетках отличаются высокой интенсивностью, сложностью и многообразием. Наряду с уже отмеченными особенностями энергетического обмена, в нервных клетках происходит синтез белков (в том числе специфических) широкого спектра, функционально активных пептидов, медиаторов и модуляторов синаптических процессов, продуктов нейросекреции. Электрические процессы имеют важнейшее значение для информационной деятельности нервных клеток и должны быть рассмотрены отдельно.
2.2. Электрические процессы в нейронах
Электрические процессы в нервных клетках включают в себя наличие постоянного потенциала покоя и медленных и быстрых изменений этого потенциала при возбуждении. Потенциал покоя является мембранным потенциалом нервной клетки и обусловлен неравномерным распределением электролитов по обе стороны клеточной мембраны. Внутри нервной клетки содержится большое количество органических анионов и катионов; в наружной среде катионов К+ при* мерно в 40 раз меньше, но высока концентрация катионов Ка+, анионов С1~. Крупные органические анионы не проникают через мембрану, а ионы К+, легко проникающие через мембрану, по закону диффузии перемещаются из области более высокой концентрации наружу. Это приводит к избытку положительных зарядов на наружной поверхности и преобладанию отрицательных зарядов на внутренней поверхности мембраны. Внутренняя поверхность мембраны заряжается отрицательно по отношению к наружной, при этом возникает электрическая сила, обеспечивающая обратное движение части ионов К* внутрь клетки, и устанавливается определенное равновесие, при котором суммарный поток ионов через мембрану будет равен нулю. Разность потенциалов между двумя сторонами мембраны при таком равновесии определяет величину мембранного потенциала. Наряду с потоками ионов Кь, являющихся основными факторами мембранного потенциала, через мембрану нервной клетки в значительно меньшем количестве движутся ионы Ма+, Са++, С1 . Они проходят через двойной ли-пидный слой мембраны по своим специальным для каждого вида ионов каналам, открывание и закрывание которых связано с изменением величины мембранного потенциала.
Влияние разницы концентраций и проницаемости основных ионов, участвующих в образовании мембранного потенциала, выражено в уравнении постоянного поля:
ЕМ = ………………………………….
Для создания разницы ионных концентраций и восполнения потерь ионов в мембране нервной клетки действует система мембранного насоса, осуществляющего активный транспорт ионов против градиента концентрации и использующего для этого энергию нейронного метаболизма. Наиболее существен натрий-калиевый насос, возвращающий К+ внутрь клетки и выводящий из нее Ма4. На внутренней стороне мембраны Ма+ соединяется с молекулой переносчика; образованный комплекс ион-переносчик проходит через мембрану; на наружной поверхности комплекс распадается, высвобождая ион N3^ и соединяясь с ионом К+, транспортирует его внутрь. Источником энергии для работы насоса служит расщепление АТФ ферментом АТФ-азой, выполняющим функцию переносчика.
Поскольку соотношение количества переносимых насосом Ма+ и К4 неодинаково, то насос не только поддерживает разницу ионных концентраций по обе стороны мембраны, но и участвует в формировании потенциала покоя, является электрогенным. Таким образом, мембранный потенциал создается в результате работы пассивных и активных механизмов, соотношение которых у разных нейронов неодинаково. Поэтому у различных нейронов величина мембранного потенциала колеблется от -80 до -40 мв, она в значительной степени зависит от особенностей его дея-
тельности и функционального состояния. При уменьшении величины мембранного потенциала покоя (деполяризации) возбудимость возрастает, при увеличении мембранного потенциала (гиперполяризации) возбудимость снижается. Возбуждение нервной клетки связано с развитием потенциала действия. Потенциал действия, или нервный импульс, представляет собой кратковременное, длящееся миллисекунды изменение мембранного потенциала, при котором уменьшается его величина, доходит до нуля и затем потенциал меняет знак. В момент пика потенциала действия мембрана становится заряженной внутри не отрицательно, а положительно (4—50 мв); амплитуда потенциала действия составляет 110—130 мв.
Перезарядка мембраны при возбуждении происходит из-за быстрого и значительного повышения мембранной проницаемости для N3% вследствие чего большое количество ионов Ыа+ проникает с наружной на внутреннюю сторону мембраны и создает здесь избыток положительных зарядов (рис. 2.2).
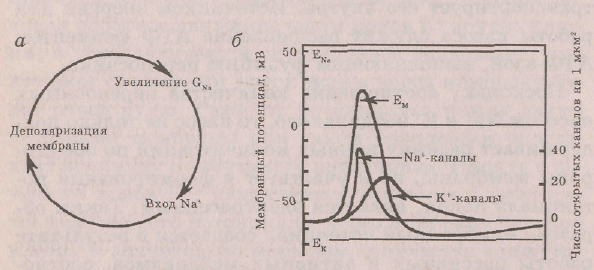
Рис. 2.2. Происхождение потенциала действия: а — связи между деполяризацией мембраны, увеличением натриевой проницаемости и входящим током Nа*; б — кривые изменения, ионных проницаемостей в процессе формирования потенциала действия (Щеперд, 1987)
Восходящая фаза потенциала действия обусловлена избирательным повышением проницаемости мембраны для Ма+. Раскрытие натриевых каналов связано с уменьшением мембранного потенциала и происходит со все возрастающей интенсивностью — лавинообразно, так как переход Ма+ на внутреннюю поверхность усиливает деполяризацию и приводит к раскрытию новых натриевых каналов. Нисходящая фаза потенциала действия связана с инактивацией натриевых каналов и повышением проницаемости для К+, так как калиевые каналы раскрываются позже натриевых.
Усиленный поток 1С наружу приводит к восстановлению мембранного потенциала до величины потенциала покоя. В телах многих нейронов потенциал действия связан и с входящим током Са++, отличающимся большей продолжительностью. Вход Са++ внутрь клетки во время потенциала действия является эффективным механизмом повышения внутриклеточной концентрации свободного Са++, который запускает или участвует в работе многих метаболических процессов. Во время возбуждения значительно усиливается работа натрий-калиевого насоса, активируемая повышением концентрации Ка+ на внутренней поверхности мембраны. Его деятельность способствует восстановлению потенциала покоя. Потенциал действия обладает порогом, при котором деполяризация достигает критического уровня и раскрываются все натриевые каналы мембраны. При под-пороговых воздействиях раскрывается лишь часть натриевых каналов, перезарядка мембраны не происходит, возникает местное возбуждение. Вследствие того, что при потенциале действия раскрываются все натриевые каналы, его амплитуда постоянна и не зависит от силы раздражения; с этим связана и невосприимчивость к новому раздражению. Потенциа-
лы действия способны быстро и надежно распространяться по мембране тела и аксона нервной клетки. Способность к распространению возбуждения связана с тем, что во время потенциала действия происходит изменение знака заряда в возбужденном участке мембраны. Между ним и невозбужденными соседними участками мембраны возникают локальные электрические токи, под действием которых происходит деполяризация новых соседних участков, что приводит к формированию в них потенциала действия. Далее развиваются локальные токи между новым участком, охваченным возбуждением, и следующими невозбужденными участками; и так возбуждение активно распространяется вдоль всей не-миелинизированной мембраны. Чем больше диаметр волокна, тем скорость распространения возбуждения выше (см. табл. 2.1).
У позвоночных большинство аксонов покрыто ми-елиновой оболочкой, периодически прерывающейся на перехватах Ранвье. В перехватах существует высокая плотность потенциалзависимых натриевых каналов (12 000 на 1 мм2), здесь генерируется потенциал действия, а на участках между перехватами возможно электротоническое формирование локальных токов, вызывающих потенциал действия лишь на следующем перехвате. Благодаря этому происходит скачкообразное (сальтаторное) распространение потенциала действия со значительно большей скоростью, чем по немиелинизированной мембране. Разновидность активного проведения возбуждения выявлена и на определенных участках дендритов некоторых нейронов.