Аминокислоты надо переносить через мембраны
Перенос аминокислот через мембраны клеток, как при всасывании из полости кишечника в энтероциты, так и при переходе из крови в клетки различных тканей, осуществляется при помощи двух механизмов: вторичный активный транспорт и глутатионовая транспортная система.
Транспорт аминокислот через мембраны Вторичный активный транспорт
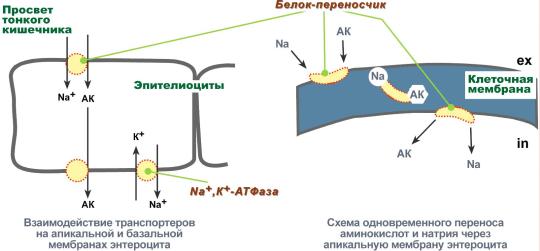
Вторичный активный транспорт – это перенос веществ, в данном случае аминокислот, с использованием градиента концентрации натрия между внутренней и наружной сторонами клеточной мембраны.
Вторичный активный транспорт основан на использовании низкой концентрации ионов натрия внутри клеток, создаваемой мембранным ферментом Na+,K+-АТФазой. Специфический белок-транспортер связывает на апикальной поверхности энтероцитов аминокислоту и ион натрия. Важно то, что в отсутствие натрия аминокислота не в состоянии связаться с белком-переносчиком.
Затем, изменив свое положение в мембране, белок отдает ион натрия в цитозоль по градиенту концентрации. Сразу после этого аминокислота теряет связь с белком и остается в цитоплазме.
Вторичный активный транспорт аминокислот через мембраны
В настоящее время выделяют 5 транспортных систем:
для крупных нейтральных, в том числе алифатических и ароматических аминокислот,
для малых нейтральных – аланина, серина, треонина,
для основных аминокислот – аргинина и лизина,
для кислых аминокислот – аспартата и глутамата,
для малых аминокислот – глицина, пролина и оксипролина.
Глутатионовая система транспорта
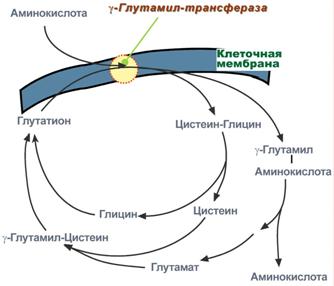
Второй способ переноса аминокислот внутрь клетки происходит в комплексе с глутатионом при помощи фермента γ-глутамилтрансферазы.
.
Транспорт аминокислот при участии глутатиона
Переносчиком некоторых аминокислот (обычно нейтральных) по этой схеме является трипептид глутатион (γ-глутамилцистеилглицин). При взаимодействии глутатиона с аминокислотой на внешней стороне клеточной мембраны при участии глутамилтрансферазы. γ-Глутамильный остаток связывает аминокислоту и происходит ее перемещение внутрь клетки. Глутатион при этом распадается на составляющие. После отделения аминокислоты происходит ресинтез глутатиона.
Аминокислоты могут давать энергию
(см. выше)
Нейромедиаторы образуются из аминокислот(см. выше)
Существуют 4 типа дезаминирования (см выше)
Катаболизм аминокислот начинается с удаления аминогруппы
У человека основным способом дезаминирования является окислительное дезаминирование. Выделяют два варианта окислительного дезаминирования: прямое и непрямое.
Прямое окислительное дезаминирование
Прямое дезаминирование катализируется одним ферментом, в результате образуется NH3 и кетокислота. Прямое окислительное дезаминирование может идти в присутствии кислорода (аэробное) и не нуждаться в кислороде (анаэробное).
1. Аэробное прямое окислительное дезаминирование катализируется оксидазами D-аминокислот (D-оксидазы) в качестве кофермента использующими ФАД, и оксидазами L-аминокислот (L-оксидазы) с коферментом ФМН. В организме человека эти ферменты присутствуют, но практически неактивны.

Реакция, катализируемая оксидазами d- и l-аминокислот
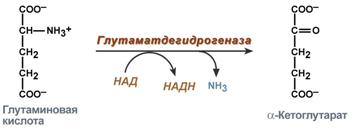
2. Анаэробное прямое окислительное дезаминирование существует только для глутаминовой кислоты, катализируется толькоглутаматдегидрогеназой, превращающей глутамат в α-кетоглутарат. Фермент глутаматдегидрогеназа имеется в митохондриях всех клеток организма (кроме мышечных). Этот тип дезаминирования теснейшим образом связан с трансаминированием аминокислот и формирует с ним процесстрансдезаминирования (см ниже).