

Билет 27.
Закончивший стометровку спринтер в начале дышит тяжело, но постепенно его дыхание выравнивается и становится нормальным. К этому периоду возвращается к норме и содержание лак. За время восстановления (до 30 мин) лактат удаляется из крови в печень и превращается в процессе гликонеогенеза в глю крови. Глю крови возвращается в мышцы:
Алкоголь тормозит глюконеогенез.
Потребление больших количеств алкоголя резко тормозит глюконеогенез в печени, вследствие чего понижается содержание глю в крови, т.е. возникает гипогликемия. Это особенно сказывается после тяжелой физической нагрузки и на голодный желудок, уровень глю может понизиться до 40 и даже 30% от нормы. Гипогликемия не благоприятно сказывается на функции мозга. Она особенно опасна для тех областей мозга, которые контролируют температуру тела. Температура тела может понизиься на 20С. Старый обычай, предписывающий давать спасенным на море или в пустыне голодным или обессилившим людям водку, физиологически неоправдан и даже опасен; в таких случаях следует давать глю.
В глюкозу могут превращаться глюкогенные аминокислоты: аланин, глутамат, аспартат, которые превращаются соответственно в пируват, оксалоацетат и -кетоглутарат:
Глюкозо-аланиновый цикл:
Глюкоза в мышцах превращается в пируват в мышцах,он превращается в аланин в мышцах,который транспортируется в пкчень,аланин в печени превращается в глюкозу,которая транспортируется в мышцы.
Данные циклы не приводят к увеличению количества глюкозы в мышцах,но они решают проблемы транспорта аминного азота из мышц в печень и предотвращают лактацидоз.
Билет 28.
Обмен гликогена.
Глюкоза как моносахарид свободно диффундирует через мембрану печеночных клеток, она не может служить резервом углеводородов в печени. Так как фосфатные эфиры глюкозы и фруктозы не столь легко проникают через мембраны, фосфорилирование этих сахаров создает как бы ловушку для них, эффективно изолируя реакции гликолиза и фосфоглюконатного пути в пределах внутриклеточного отсека. Тем не менее фосфаты сахаров не могут накапливаться в больших количествах в печени, так как благодаря осмосу увеличение их запасов будет сопровождаться накоплением больших количеств воды.
В печени происходит превращение избытка углеводов в нерастворимый полимер – гликоген. Эта резервная форма глюкозы может составлять одну десятую всей массы печени. При голодании запас гликогена почти полностью истощается.
Обмен гликогена включает в себя 2 процесса: гликогенез или синтез гликогена и гликогенолиз или распад гликогена.
Гликогенез – синтез гликогена – это анаболический процесс, требующий затраты энергии в форме как АТФ, так и уридинтрифосфата (УТФ). Исходным источником для синтеза гликогена является глю, d под действием глюкокиназы превращается в глю-6-фосфат.
Глю-6-ф занимает ключевое положение как общий промежуточный продукт ряда процессов углеводного обмена: он способен к обратимому превращению в пируват путем гликолиза или глюконеогенеза, а также к необратимому – в пентозы в ходе фосфоглюконатного пути. Глю-6-ф может обратимо превращаться в гликоген.
Затем глю-6-ф превращается в глю-1-ф. В процессе активации глю-1-ф участвуют уридиннуклеотиды. В ходе этого процесса два концевых фосфорных остатка УДФ освобождаются в виде неорганического пирофосфата, остаток УМФ соединяется с глю-1-ф, образуя УДФ-глюкозу.
Источником глю при синтезе гликогена является УДФ-глю, d образуется из глюкозо-1-ф и УТФ:
Глю-1-ф + УТФ
![]() УДФ=глю
+ Н4Р2О7
УДФ=глю
+ Н4Р2О7
На следующей стадии происходит перенос остатка глю с УДФ-глю на затравку гликогена с помощью гликогенсинтетазы:
УДФ-глю + (глю)n УДФ +(глю)n+1
Гликогенсинтетаза катализирует образование только -1,4-гликозидных связей. «Ветвящий» фермент образует -1-6 гликозидные S (точки ветвления).
Гликогенолиз – распад гликогена идет путем фосфоролиза.
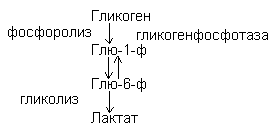
Гормональная регуляция.
инсулина
В печени активирует гликогенсинтетазу, усиливает синтез гликогена – гликогенез.
Инсулин активирует:
Ферменты гликогенеза (синтез гликогена): гликогенсинтазу.
Механизм действия адреналина и глюкагона
Усиливают распад гликогена в мышцах и печени, активируя фосфорилазу гликогена и переход глюкозы в кровь за счет активизации глюкозо-6-фосфотазы. Адреналин оказывает преимущественное действие на мышечные клетки, а глюкагон – на клетки печени.
Гормрнальная регуляция обмена глюкозы.
Билет 30.
Эффект Пастера.
Снижение скорости потребления глюкозы и прекращение накопления лактата в присутствие кислорода осит название эффекта Пастера.Впервые это явление наблюдал Л Пастер во время своих широко известных исследований болезней вина.В дальнейшем было показано,что эффект Пастераа наблюдается также в животных и растительных тканях,где кислород тормозит анаэробный гликолиз .Значение эффекта Пастера ,т е перехода в присутствие кислорода от анаэробного гликолиза,или брожения ,к дыханию,состоит в переключении клетки на наиболее эффективный и экономичный пут получения энергии.В результате скороть потребления субстрата в присутствие кислорода снижается.Молекулярный механизм эффекта Пастера заключается,по-видимому,в конкуренции между системами дыхания и гликолиза за АДФ,используемый для образования АТФ. Как известно в аэробных условиях значительно эффективнее,чем в анаэробных,происходят удаление Ф и АДФ,генрация АТФ,а также регенерирование НАД+ из НАДН.Т о уменьшение в присутствие кислорода количества Адф и Ф и соответствующее увеличение количества АТФ ведут к подавлению анаэробного гликолиза.
Регуляция обмена углеводов.
Соотношение между процессами катаболизма и анаболизма глюкозы в клетках печени находятся под контролем целого ряда факторов регуляции:
Концентрация метаболитов и глюкозы.
Воздействие гормонов.
Внутриклеточные рецепторы.
Субстраты и промежуточные продукты метаболических реакций играют важную роль в регуляции соотношения между гликолизом и глюконеогенезом.
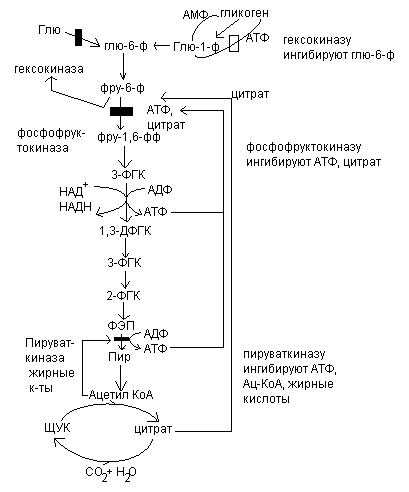
Рассмотрим, как регулируется само вступление остатков глюкозы на путь гликолиза. Вовлечение глюкозных остатков в процесс гликолиза обеспечивает важная реакция и эта реакция контролируется регуляторным ферментом. Реакция катализируется гексокиназой и активность этого фермента ингибируется глюкозо-6-фосфатом. Когда концентрация глюкозо-6-фосфата в клетке сильно возрастает, т.е. когда он образуется быстрее, чем потребляется, наступает ингибирование – гексокиназа под действием глюкозо-6-фосфата выключается и дальнейшего фосфорилирования глюкозы не происходит до тех пор, пока избыток глюкозо-6-фосфата не будет использован.
Г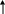
 лю
лю
![]() Глю-6-ф
Глю-6-ф
 В
печени преобладает другой фермент –
глюкокиназа, которая не ингибируется
глюкозо-6-фосфатом. Поэтому в печени,
способной хранить большие количества
гликогена, избыточная глюкоза крови
может фосфорилироваться в глюкозо-6-фосфат,
который через глюкозо-1-фосфат превращается
в гликоген.
В
печени преобладает другой фермент –
глюкокиназа, которая не ингибируется
глюкозо-6-фосфатом. Поэтому в печени,
способной хранить большие количества
гликогена, избыточная глюкоза крови
может фосфорилироваться в глюкозо-6-фосфат,
который через глюкозо-1-фосфат превращается
в гликоген.
Кроме гексокиназной реакции в гликолизе имеются еще два главных регулируемых этапа: это фосфофруктокиназная и пируваткиназная реакция.
В скелетных мышцах активность фосфофруктокиназы определяется концентрациями субстратов этого фермента (АТФ и фруктозо-6-фосфата) и его продуктов (АДФ и фруктозо-1,6-дифосфата), а также цитрат.
Главными отрицательными модуляторами фосфофруктокиназой являются АТФ и цитрат.
Главными положительными модуляторами являются АМФ и фруктозо-1,6-дифосфат.
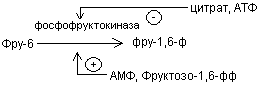
Третьим регулируемым этапом гликолиза является пируваткиназная реакция.
![]()
Активность пируваткиназы ингибируется АТФ и ацетил-КоА, а также жирные кислоты.
Реакция |
Фермент |
Ингибитор |
Активатор |
Глю Глю-6-ф |
Глю-6-ф |
|
|
Фру-6-ф |
АТФ, цитрат, ацил-КоА, кетоновые тела, НАДН+Н+ |
Фру-1,6-дф, АДФ, НАД |
|
ФЕП |
АТФ, ацетил-КоА, жирные к-ты, аланин |
АДФ, Фн, НАД, фру-1,6-дф |
|
Пир |
|
Ацил-КоА, Ацетил-КоА, Аланин |
|
ЩУК |
|
Ацил-КоА, Ацетил-КоА, Аланин |
|
Регуляция гликолиза.
Три стадии катаболизма углеводов обеспечивают получение энергии: гликолиз, цикл лимонной кислоты и окислительное фосфорилирование.
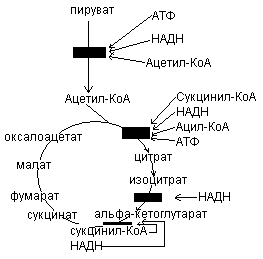
При уменьшении содержания АТФ и Рн, это приводит к возрастанию скорости переноса электронов и окислительного фосфорилирования. Одновременно повышается скорость окисления пирувата через цикл лимонной кислоты, т.е. усиливается приток электронов в дыхательную цепь. Это приводит к увеличению скорости гликолиза, тем самым обеспечивается образование пирувата. Затем наступает момент, когда отношение АТФ/АДФ+Рн возвращается к обычному высокому уровню. Теперь перенос электронов и окислительное фосфорилирование замедляются. Цикл лимонной кислоты замедляется, т.к. АТФ и цитрат являются ингибитором фосфофруктокиназы и пируваткиназы.
Регуляция пируватдегидрогеназного комплекса.
Превращение пирувата в Ацетил-КоА происходит при участии пируватдегидрогеназного комплекса, содержащего 5 коферментов: НАД, ФАД, НSКоА, ТДФ, липоевую кислоту. В основе процесса лежит реакция окислительного декарбоксилированя.
Образование ацетил-КоА из пирувата – это ключевой необратимый этап метаболизма, потому что животные неспособны к превращению ацетил-КоА в глюкозу.
Окислительной декарбоксилирование пирувата в ацетил-КоА создает возможность превращения атомов глюкозы по 2 путям: 1) окисление до СО2 в ЦТК с одновременным регенерированием энергии; 2) включение в липиды. Это позволяет считать, что активность пируватдегидрогеназного комплекса должна строго регулироваться.
Активность ПДК (пируватдегидрогеназного комплекса) регулируется 3 путями:
Ингибирование продуктами реакции. Ацетил-КоА и НАДН ингибируют превращение

Регуляция нуклеотидами по принципу обратной связи.
Пируватдегидрогеназный комплекс ингибируется ГТФ и активируется АМФ.
Активность комплекса снижается, когда клетка богата легкодоступной энергией.
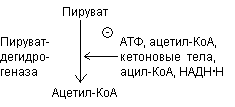
Регуляция путем ковалентной модификации. Комплекс теряет ферментативную активность, когда специфический остаток серина пируватдегидрогеназного комплекса фосфорилируется АТФ. Фосфорилирование усиливается при высоких соотношениях АТФ\АДФ, ацетил-КоА\СоА, НАДН\НАД+ и ингибируется пируватом.
Регуляция ЦТК.
Скорость
функционирования ЦТК зависит от
потребности клетки в АТФ. Важной
регуляторной реакцией цикла является
синтез цитрата из ацетил-КоА и оксалоацетата
под действием цитратсинтетазы. АТФ –
аллостерический ингибитор цитратсинтетазы.
1) Ац-КоА + ЩУК![]() цитрат
цитрат
Вторая регуляторная реакция – это реакция, катализируемая изоцитрат-дегидрогеназой. Фермент аллостерически активируется АДФ, НАДН ингибирует изоцитрат-дегидрогеназу:
2) Изоцитрат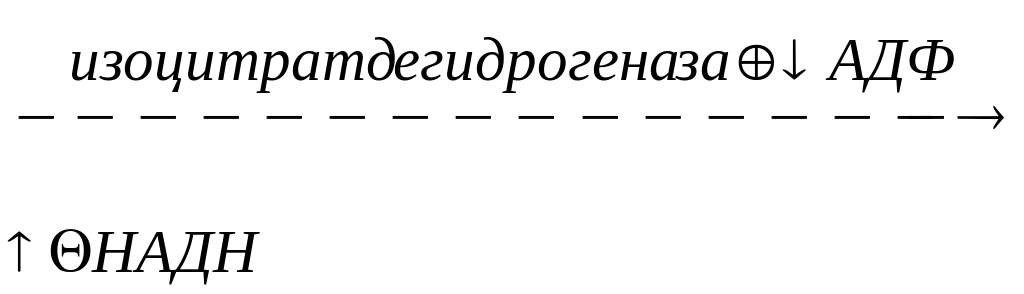 -оксоглутарат
-оксоглутарат
3) Третьей регуляторной реакцией является реакция, катализируемая -кетоглутаратдегидрогеназой:
-кетоглутарат![]() сукцинил-КоА
сукцинил-КоА
оксоглутарат
Регуляция цикла лимонной кислоты.
Активация пируваткарбоксилазы.
Активность пируваткарбоксилазы зависит от присутствия ацетил-КоА.
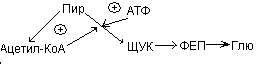
Пир может превращаться в ацетил-КоА, а также подвергаться карбоксилированию с образование оксалоацетата или ЩУК. Высокое содержание ацетил-КоА приводит к образованию количества оксалоацетата. Если имеет место избыток АТФ, то оксалоацетат потребляется в процессе глюконеогенеза:
Ацетил-КоА![]() ЩУК
ЩУК![]() ФЕП
Глю
ФЕП
Глю
В условиях недостатка АТФ оксалоацетат включается в ЦТК, конденсируясь с ацетил-КоА.

Гормональная регуляция обмена углеводов
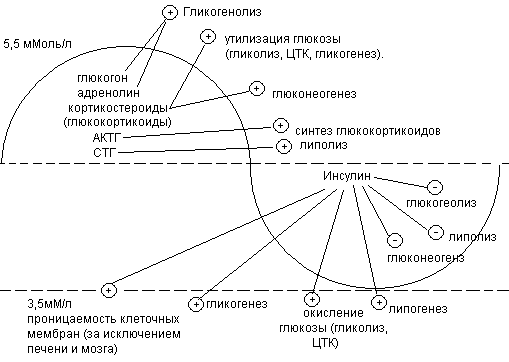
Основным показателем состояния углеводного обмена является содержание глюкозы в крови. В норме содержание глюкозы составляет 3,5 – 5,5 ммоль/л.
Снижение содержания глюкозы ниже 3,3 ммоль/л называется гипогликемия. При снижении содержания глюкозы ниже 2,7 ммоль/л развивается грозное осложнение – гипогликемическая кома. Содержание глюкозы в крови выше 6 ммоль/л называется гипергликемией. Если содержание глюкозы превышает 50 ммоль/л, развивается гипергликемическая кома. При увеличении содержания глюкозы в крови выше 10 ммоль/л глюкоза появляется в моче и возникает глюкозурия.
Инсулин – единственный гормон гипогликемического действия (снижает уровень глюкозы).
Адреналин, клюкагон, АКТГ, СТГ, глюкокортикоиды – гипергликемические гормоны(повышают уровень глюкозы).
Механизм действия инсулина
Повышает проницаемость клеточных мембран для глюкозы, способствуя переходу ее из крови в ткани;
задерживает глюкозу в клетках, активируя гексокиназу («гексокиназная ловушка глюкозы»);
Усиливает распад глюкозы в мышцах путем индукции синтеза регуляторных ферментов гликолиза – гексокиназы, фосфофруктокиназы, пируваткиназы;
В печени активирует гликогенсинтетазу, усиливает синтез гликогена – гликогенез.
Подавляет синтез ферментов глюконеогенеза, препятствует избыточному катаболизму жиров и белков и переходу их в углеводы.
Инсулин регулирует активность ферментов на генетическом уровне – является индуктором синтеза ферментов гликолиза и репрессором синтеза ферментов глюконеогенеза.
Инсулин активирует дегидрогеназы пентофосфатного пути.
Инсулин активирует:
Ферменты гликолиза: гексокиназу, фосфофруктокиназу, пируваткиназу.
Ферменты пентозофосфатного пути: глюкозо-6-фосфатдегидрогеназу, 6-фосфоглюконатдегидрогеназу.
Ферменты гликогенеза (синтез гликогена): гликогенсинтазу.
Ферменты ЦТК: цитратсинтазу.
Механизм действия адреналина и глюкагона
Усиливают распад гликогена в мышцах и печени, активируя фосфорилазу гликогена и переход глюкозы в кровь за счет активизации глюкозо-6-фосфотазы. Адреналин оказывает преимущественное действие на мышечные клетки, а глюкагон – на клетки печени.
Механизм действия глюкокортикоидов.
Усиливают глюкогенез за счет индукции синтеза в клетках печени ключевых ферментов глюкогенеза – фосфоенолпируват-карбоксилазы, пируваткарбоксилазы, фруктозо-1,6-дифосфотазы, глюкозо-6-фосфотазы.
Гормрнальная регуляция обмена глюкозы.
Механизм действия гормонов заключается в повышении (снижении) активности готовых форм ферментов или (глюкокортикоиды) + интенсификация их синтеза.
Гипергликемические гормоны:
Адреналин, глюкогон – активация фосфорилазы.
К![]() ортикостероиды
– активация (усиление синтеза) ферментов
глюконеогенеза: пируваткарбоксилаза,
ФЕП-карбоксилаза, фру-1,6-дифосфотаза,
глю-6-фосфотаза.
ортикостероиды
– активация (усиление синтеза) ферментов
глюконеогенеза: пируваткарбоксилаза,
ФЕП-карбоксилаза, фру-1,6-дифосфотаза,
глю-6-фосфотаза.
- утилизация глюкозы – ингибируют гексокиназу
АКТГ - усиление синтеза гормонов коры надпочечников
СТГ – опосредованное действие, активируя липазу жировых депо и способствуя повышению концентрации НЭЖК в крови (энергетический материал), сберегается глюкоза.