

Генетическая инженерия и безопасность
.pdf11
сраз у в клетку эукариотили некоторы х б актерий (б ез вектора)? Э то свя з а- но с тем, ч то при об ы ч ном введении Д Н К в клетку она, какправило, подвергается атакеф ерментов, которы еразрезаю теенаотдельны еф рагменты . Д ля того, ч тоб ы рекД Н К сталасос тавной ч астью генетич ес кого аппарата клетки, онадолжналиб о встроиться в ее геном (интегрировать в хромосому) и реплицироватьс я з аего сч ет, либ о б ы ть с пособ ной кавтономной репликации.
В ектордолженоб ладать с ледую щ ими свой ствами.
1. |
С пособ ность к автономной (т.е. нез ависимо отхромосомы реци- |
пиента) |
репликации в клетке реципиента. Н апример, для репликации в |
клетке б актерии, вектор должен с одержать с ай т ori (уч ас ток инициации репликации).
2. Н алич ие с ай та, в котором возможно вс траивание желаемого ф рагментаД Н К . Д ля этоговектордолженсодержать один, или самоеб ольш оедвауч астка(сай тарес трикции), ч увствительны х к определенной рестриктазе, которая рас щ епля етвектори поз воля етвс троить желаемы й трансген.
3. Н алич ие одного или нескольких маркерны х генов, б лагодаря которы м клетка-реципиент б удет об ладать новы ми признаками, позволяю - щ ими отлич ить трансф ормированны е клетки (т.е. содержащ ие рекД Н К ) от исходны х. Э то могут б ы ть селект и вн ы еген ы , которы е придаю т клеткам селективное преимущ ес тво (ус той ч ивость к антиб иотикам, герб ицидам). Такие гены кодирую тф ерменты , разруш аю щ ие или модиф ицирую щ ие антиб иотики, герб ициды . В этом с луч ае трансф орманты отб ираю т на питательны х средах с вы соким содержанием э тих вещ еств. Н апример, в присутствии геналактомаз ы б актериальная клеткаприоб ретаетустой ч ивость к пенициллину и на с реде с этим антиб иотиком об разуетклон (нес ущ ий данны й ген), тогдакак об ы ч ны е клетки (б ез этого гена) на данной среде погиб аю т. В кач естве маркерны х использ ую ти такназы ваемы е р еп ор т ер - н ы егены , экспресс ия которы х не даетс елективны х преимущ еств, но продукты генов удоб ны для тестирования , например, по из менению окрас ки. Так, ген GFP контролируетс интез з еленого ф лю орес цирую щ его б елкаиз
медузы . П ри об луч ении транс генны х растений , |
с одержащ их этот б елок, |
У Ф -луч ами, поя вля ется зеленое свеч ение. Г ены |
luxA и luxB, вы деляю тиз |
Д Н К светля ч ков. О ни контролирую тс интез лю циф ераз ы , которая об еспе- ч ивает переход лю цеф иринов из окисленной ф ормы в основную , ч то и об еспеч иваетсвеч ение трансгенны х растений , накапливаю щ их этотб елок.
Ш ироко использ уемы м в настоя щ ее время репортерны м геном |
я вля ется |
ген β-глю коронидазы (GUS). Трансгенны е клетки, экспрес сирую |
щ ие этот |
ген, при помещ ении их нас пециф ич еский суб страт, |
окраш иваю тся в голу- |
б ой цвет. |
|
4. Кроме того, для того ч тоб ы ч ужеродны й |
ген экспресс ировалс я , |
необ ходимо его помес тить под с оответствую щ ий промотор. У эукариоти- ч еских организ мов механизм регуля ции транскрипции б олеес ложны й , ч ем у эукариот. Регуля торны е пос ледовательности э укариотич еских генов отлич аю тся отпрокариотич еских, и б актериальная РН К -полимеразане уз на-
12
етих. П оэтому для э кс прессии эукариотич еских генов в клетках прокариот нужно, ч тоб ы гены находилис ь под контролем б актериального промотора (т.е. промотораклетки-хозя ина). В кач естве промотораш ироко ис пользуется промотор генаβ-лактомаз ы (ген ус той ч ивости кампициллину), локализ ованногов векторерBR322, lac-промоторE. coli и др.
То ес ть соз дается целая генетич еская конс трукция , в состав которой , помимо трансгена, вводя тся маркерны е гены и с оответствую щ ие регуля - торны епос ледовательности.
В кач естве векторны х молекул могутб ы ть использованы плаз миды б актерий или дрожжей (просты х э укариотич еских организ мов), Д Н К б ак-
териоф агов или вирусов, искусс твенны е хромосомы дрожжей |
(YAK) и |
б актерий (BAK). С озданы также гиб ридны е (ис кус ственны е) |
векторы - |
кос миды , об ъ единя ю щ иепреимущ естваплазмид и ф агов. |
|
Плазми ды – внехромосомны е генетич еские элементы про- и эукариот, которы е автономно реплицирую тся в клетке. П риродны е плаз миды ч ас то с одержатгены , полез ны е для б актерий : придаю щ ие устой ч ивость к антиб иотикам, контролирую щ ие способ ность разруш ать раз лич ны е труднораз лагаемы е токс ич еские соединения (наф талин, камф ору, толуол, ксилол, различ ны е пес тициды и др.). Благодаря этому, например, б актерии родаPseudomonas сущ ес твую тв различ ны х экологич еских ниш ах, в неб ла-
гоприя тны х |
ус ловия х окружаю щ ей |
среды , их ис пользую т для оч истки |
поч вы , воды |
и з агря з нений токс ич |
ес кими соединения ми. С плаз мидами |
свя з анаспособ ность ря дапоч венны х б актерий вс тупать в симб иоз с б об о-
вы ми растения ми, об условливая их с пос об ность к об разованию |
корневы х |
клуб еньков, необ ходимы х для ус воения поч венного азота. Д ля |
геннной |
инженерии б ольш ой интерес представля ю тмногокопий ны е (мультикопий -
ны е) плаз миды , |
которы е в клетке представлены б ольш им |
ч ис лом копий |
(до 10-200 копий |
на клетку). И с пользуя их, можно достич |
ь сверхсинтез а |
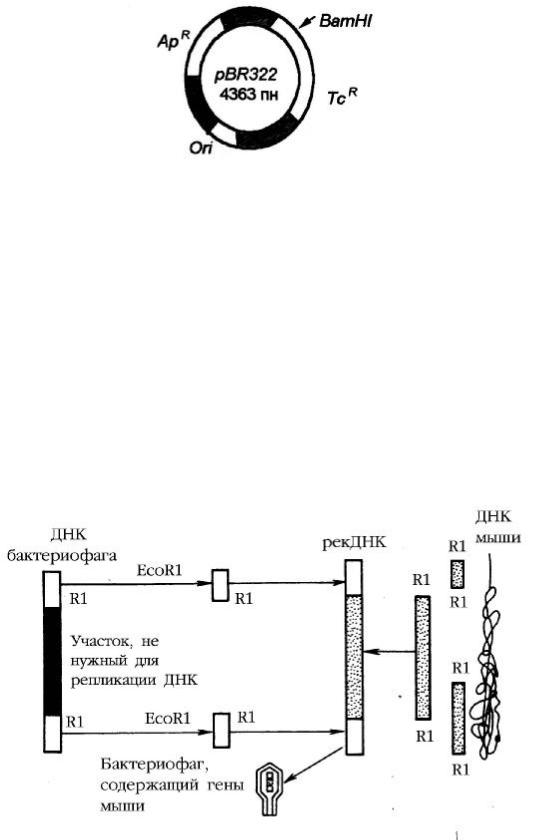
нужны х б елковы х продуктов. Ч ащ евсеговекторы конструирую тнаоснове природны х плазмид, удаля я целы й ря д лиш них генов. Н апример, ш ироко использ уемы й для этих целей плаз мидны й векторр BR322 соз даннаоснове плазмиды E.coli. П лаз мидар BR322 имеетс ай тori (об ласть, ответственную зарепликацию плаз миды ), гены устой ч ивости кантиб иотикам ампициллину (Ap′) и тетрациклину (Tc′). В генеTC′ имеетс я уникальны й сай т, разрезаемы й рестриктазой Bam HI (рис унок2). Д ля растений ис пользую тся векторы , сконструированны е наос нове Ti- и Ri- плаз мид поч венны х агроб актерий (рис унок10). Э ти б актерии поражаю тдо 60% двудольны х растений и некоторы е однодольны е растения , вы зы вая ф ормирование опухолей – коронч аты х галлов (Agrobacterium tumefaciens) или об разование “косматы х”корней (A. rhizogenes). В плаз мидах можно клонировать ф рагменты Д Н К раз мером неб олее10 тпн.
Ф аговы евект ор ы , ч ащ е всего, с оздаю т на б аз е умеренного б актериоф агаλ, содержащ егодвухцепоч еч ную линей ную молеклулД Н К (рис у - нок3). Л евоеи правоеплеч и ф агаимею твсегены , необ ходимы едля
13
Рисунок2. С хемастроения плазмиды pBR322
С елективны е маркерны е гены , определяю щ ие устой ч ивость кантиб иотикам: ампициллину (ApR) и тетрациклину (TcR); В гене TcR имеетс я уникальны й сай т, разрезаемы й рестриктазой BamHI; Ori – уч асток Д Н К, ответственны й з а репликацию плазмиды в клетках E. coli.
литич еского цикла (репликации, раз множения ). С редня я же ч асть генома б актериоф ага λ (с одержащ ая гены , контролирую щ ие лиз огению , т.е. его интеграцию в Д Н К б актерии-хозя ина) не с ущ ественнадля его размножения и около 50% (≈25тпн) можетб ы ть замененанач ужеродны й ф рагмент Д Н К. Такие модиф ицированны е ф аги проходя тлитич еский цикл, но лизогения не происходит. В екторы наоснове б актериоф агаλ использ ую тдля клонирования ф рагментов Д Н К э укариот(т.е. б олеекрупны х генов) размером до 23 тпн. П рич ем, ф аги б ез вставок (< 38 тпн) или, напротив, со слиш ком б ольш ими вставками (> 52 тпн) не развиваю тся и не поражаю т б актерии.
Рисунок3. С труктуравектора, созданногонаос новеД Н К б актериоф агаλ [Ш евелуха, 2003]
14
Косми да – это векторная плазмида, предназнач енная для клонирования б ольш их ф рагментов Д Н К эукариот(до 45тпн) в клетках E. coli. Термин об оз нач ает, ч то вектор я вля ется плазмидой , внутри которой вставлен
cos-уч асток ф ага λ (cos-sites), |
предс тавля ю щ ий соб ой нуклеотидную по- |
следовательнос ть, отвеч аю щ ую |
з аупаковку ф аговой Д Н К в ее протеино- |
вую капс улу (рисунок4). Как следс твие и плазмидная Д Н К , вклю ч аю щ ая ч ужеродны е гены , можетб ы ть упакованав космидах в протеиновую капсулу б актериоф ага.
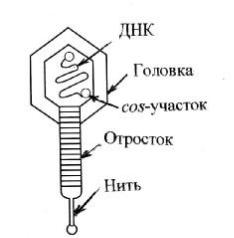
Рис унок4. С хемастроения зрелогоб актериоф агаl.
Л иней ная двухцепоч еч ная Д Н К б актериоф ага λ состоитпримерно из 50 тпн. Н а концах Д Н К имею тс я cos-уч ас тки, отвеч аю щ ую з а упаковку ф аговой Д Н К в еепротеиновую капс улу.
Зач ас тую полноразмерны е гены и мультигенны е комплекс ы (³100тпн) эукариотс лиш ком велики для встраивания в об ы ч ны е векторы . Д ля перенос а крупны х трансгенов и их клонирования использ ую т и скус-
ст вен н ы ехр омосом ы др ож ж ей (YAK- я ки отангл. yeast artificial chromosomes), вмещ аю щ ие ф рагменты геномной Д Н К длиной от100 тпндо1 млн пн. Д ля их с оз дания к плаз миде дрожжей “приш иваю т”центромерны е (CEN) последовательности, теломеры (концевы е последовательности), последовательнос ти для автономной репликации (ARS) в дрожжевой клетке, сай ты рестрикции и селективны е маркеры (TRPI и URA3 - нез авис имос ть отналич ия триптоф анаи урациласоответственно).
Ч елн очн ы е вект ор ы . Э то векторы (сконструированны е на основе плазмидной Д Н К ), с пособ ны е реплицироваться в клетках двух и б олее ор-
ганиз мов. |
Н апример, плазмида YEp24 с пособ на раз множатьс я в клетках |
дрожжей |
и E. coli. В э том случ ае векторы имею тс пециф ич еские нуклео- |
тидны е последовательности (специф ич ны е для дрожжей и E. coli), поз воля ю щ ие реплицироваться или в б актерии, или в дрожжевой клетке. С помощ ью ч елноч ного вектораудалос ь ввес ти гены лей коцитарного интерф е- ронач еловекав клетки дрожжей . С конструирован ш тамм дрожжей , кото-
ры й вы деля ет |
в культуральную среду поч ти ч исты е a-, b- и g- |
интерф ероны . И |
нтерф ерон– ценны й лекарственны й препарат, ш ироко ис – |
15
пользуемы й для б орьб ы с вирус ны ми инф екция ми и другими заб олевания ми, вклю ч ая з локач ественны еопухоли.
Типич ная с хемаопы тапо генетич еской инженерии предс тавленана
рис унках 5 и 6. Э кс периментвклю ч аетследую щ иеэтапы . |
|
|
1,2. |
Д ля конс труирования рекомб инантной Д Н К (рекД Н К) |
вектор- |
ную Д Н К |
(например, плазмиду) и ч ужеродную Д Н К , содержащ ую |
интере- |
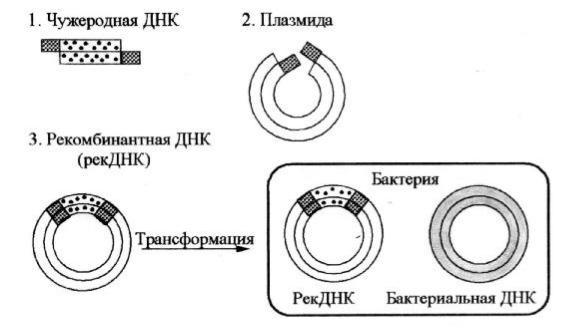
сую щ ий нас ген (трансген), разрез аю тодной и той же рестриктазой . О б разую тся одинаковы е “липкие”концы (рисунок 5). К генам, синтез ированны м химич еским путем или получ енны м по матрицеих мРН К , такие “липкие”концы можноприш ить искусственно.
3. С меш ивание раз лич ны х по происхождению ф рагментов Д Н К и сш ивание их Д Н К-лигазой . Л ипкие концы ч ужеродной Д Н К и плаз миды вз аимодей с твую т друг с другом, об разуя комплементарны е пары основа-
ний . П роисходит гиб ридиз ация |
векторной и ч ужеродной Д Н К . “Л ипкие” |
концы з амы каю тс я с помощ ью |
водородны х свя з ей , аковалентны е сш ива- |
ютс помощ ью ф ерментаД Н К -лигазы .
4.Г енетич еская трансф ормация , т.е. перенос и вклю ч ение рекД Н К,
содержащ ей транс ген, в клетки реципиента(например, E. coli). П лазмида, вс троенная в б актерию , ведет с еб я как вектор (переносч ик) нового гена, которы й реплицируетс я в каждом новом поколении.
5. М олекулярная селекция – отб ор трансф ормантов, т.е. клонов, несущ их рекД Н К. В процесс е генетич еской транс ф ормации E. coli могутоб - разоваться 3 типаклеток: не содержащ ие пламиду, с одержащ ие плаз миду б ез вс трой ки (б ез рекД Н К), с одержащ ие плаз миду с рекД Н К. Д ля отб ора трансф ормантов с реди нетранс ф ормированны х клетокиспольз ую траз лич - ны е маркерны е гены , которы е находя тся в векторной молекуле наря ду с транс геном.
Рисунок5. С хемаопы тапогенетич еской инженерии (конструирование рекД Н К )
16
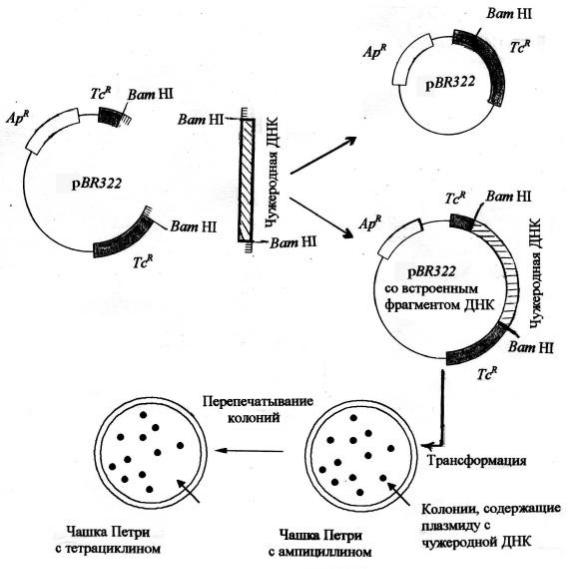
Так, плаз мидаpBR322 имеетдвагенаус той ч ивости к антиб иотикам ампициллину (ApR) и тетрациклину (TcR). О дин из них с лужитдля иденти- ф икации б актерий , несущ их плазмиду (вектор) путем отб ора клеток, ус - той ч ивы х к антиб иотику, а другой – для отлич ия гиб ридной плаз миды (рекД Н К) отродительского вектора. В гене TcR имеетс я уникальны й сай т, разрезаемы й рестриктаз ой BamHI (рисунок6). П редположим, мы разрез а-
Рис унок6. С хемаинтеграции ч ужеродной Д Н К в плаз миду pBRR322 и отб ортрансф ормированны х клонов E. coli, содержащ их плазмиду с рекД Н К [Ай ала, 1987], (поя снениев тексте)
ли вектор в гене TcR рес триктазой BamHI и встроили в него ф рагментч у-
жеродной Д Н К , |
получ енны й при помощ и той же рестриктазы . |
Г ен TcR |
инактивируетс я, |
следовательно, у б актерий , несущ их плазмиду, |
исч ез ает |
ус той ч ивос ть к тетрациклину, но сохраня ется устой ч ивость к ампициллину. О тб орнасреде с ампициллином покажет, содержитли E. coli плаз миду или нет. С одержащ ие плазмиду б актерии б удутрасти насреде с ампициллином. Д ля отб ораклеток, нес ущ их ч ужеродную Д Н К (интересую щ ий нас
17
ген), б актерии вы ращ иваю тнасреде с тетрациклином. Трансф ормированны е клетки устой ч ивы к ампициллину, но ч увствительны к тетрациклину (такие колонии отсутствую тнасреде с тетрациклином), т.к. ген устой ч и- вости к тетрациклину раз руш ен в рез ультате инс ерции ф рагмента ч ужеродной Д Н К . С трелкой нарисунке 6 отмеч ены колонии - транс ф орманты , которы есоб ираю тдля опы тов с ампициллиновогогазона.
|
1.3.2. М ет оды п ереноса геноввклет ки разли чны хоргани зм ов |
|
И з вестны многоч исленны е методы , с помощ ью которы х можно вне- |
дрить ч ужеродную Д Н К в геном различ ны х организ мов. |
|
|
В кач естве реципиентов, в геном которы х встраиваю тс я ч ужеродны е |
гены , |
ис пользую т клетки культуры , эмб риональны е клетки млекопитаю - |
щ их, |
некоторы х растений , дрозоф илы , пронуклеус ы млекопитаю щ их, у |
растений – протоплас ты , из олированны е клетки и ткани, микрос поры , незрелы е з иготич еские зароды ш и, проростки. Трансф ормация э кзогенной Д Н К можетос ущ ествля ться либ о в культуреклеток(in vitro=ex vivo), либ о непосредственнов организме(in vivo).
- М икроинъ екция. С помощ ью тонких микроигл и микроманипуля - торав клетку или пря мо в я дро вводится векторная Д Н К с вклю ч енны м в нее трансгеном. С помощ ью микроинъ екций осущ ествля ется трансф ормация у дрозоф илы , рас тений .
- Э лектропорация . Рас тительны е протоплас ты или животны е клетки об раб аты ваю тимпульс ами э лектрич еского поля вы сокого напря жения , ч то об ратимо увелич ивает проницаемость б иомемб ран. Ч ерез об раз ую щ иеся накороткоевремя поры ч ужеродная Д Н К проникаетв клетку.
- П еренос Д Н К в сос таве липосом. Л ипосомы – э то искусс твенно соз данны е сф ерич еские об разования , об олоч какоторы х состоитиз ф ос ф о- липидов. Л ипосомы , содержащ ие внутри транс ф ормирую щ ую Д Н К, способ ны непосредственно сливатьс я с мемб раной клетки или поглощ аться клетками в рез ультате процес са, подоб ного эндоцитоз у. В клетке происходитразруш ение об олоч ки липосом и вы с воб ождение рекД Н К. Э тоодиниз методов, использ уемы й для з ащ иты трансф ормирую щ его генетич еского материала от разруш ительного дей ствия нуклеаз, присутствую щ их вне клеток. М етод применя етс я для введения нуклеиновы х кислотв культивируемы еживотны еклетки, растительны епротопласты .
- Бомб ардировкамикропуля ми (б аллистич ес кая трансф ормация ). Э то один из самы х э ф ф ективны х методов транс ф ормации однодольны х и хвой ны х рас тений (в которы е не удается ввес ти ч ужеродную Д Н К с помо- щ ью агроб актерий ), атакже трансф ормации животны х клеток. Таким путем проводятгенотерапию (т.е. ис правление наследс твенны х деф ектов путем введения в геном полноценны х генов) у животны х и ч еловека. Д ля “об стрела”тканей ис пользую тс я ч астицы из з олотаили вольф рамаразмером 0,6-3 мкм, накоторы е нанос итс я Д Н К вектора, содержащ ий трансген. Э тими ч астицами (“микропуля ми”) заряжаю т“генны е”пуш ки. М икропули раз гоня ю тс я в установке под дей с твием э лектрич еского разря да или под
18
давлением газ агелия . П ри достаточ ной скорости эти ч астицы могутнепосредс твенно проникать в я дро, ч то с ильно повы ш ает э ф ф ективность трансф ормации. Э тим же методом можно трансф ормировать и другие Д Н К -содержащ иеорганеллы – хлоропласты и митохондрии.
Д ля многих двудольны х растений э ф ф ективнавекторная трансф ормация наоснове Ti- и Ri- плазмид с помощ ью агроб актерий . Э ф ф ективны ми перенос ч иками Д Н К в клетки млекопитаю щ их я вля ю тся “природны е ш прицы ”– вирус ы .
1.3.3. Клони ровани е генов |
|
Клонирование генов проводя т с |
целью получ ения того или иного |
ф рагментаД Н К в б ольш ом колич ес тве. |
Э тотпроцесс необ ходим для по- |
луч ения многоч исленны х копий желаемы х генов. Клонирование Д Н К воз - можноб лагодаря способ нос ти б актериальны х плазмид и ф агов продолжать нормальное ф ункционирование после встраивания в их геном ч ужеродной
Д Н К. |
П ос кольку встроенны е в геном ч ужеродны е последовательнос ти |
Д Н К |
не влия ю т на с вой с тва химерны х ш таммов б актерий , практич ески |
лю б ая последовательность Д Н К можетб ы ть клонированатаким об разом. Д ля клонирования плазмиду, содержащ ую рекД Н К, вводятв клетки б актерии (например, E. coli) или дрожжей , где происходит ее многократная репликация . Д ля клонирования неб ольш их ф рагментов Д Н К ис пользую т плазмиды , ф аговы е Д Н К, адля крупны х - космиды и искусственны е хромосомы . Клонирование рекомб инантны х молекул с генами ч еловека или животны х в клетках б актерий даетвозможность в ус ловиях микроб иологич ес кого синтез аполуч ать б ольш ое колич ество нужны х б елков. Так, ис - кус ственно синтез ированны й ч еловеч еский ген инс улина введен в б актерию , ч то дало возможность получ ать ч еловеч еский инс улин (гормон, ш и- роко ис пользуемы й в медицине при леч ении с ахарного диаб ета) в промы ш ленны х колич ествах. Раньш е для леч ения с ахарного диаб етаис пользовали инсулин животного происхождения , получ аемы й из поджелудоч - ной желез ы крупного рогатого скота. В 1979 г. из 60 млн. б ольны х диаб е- том во вс ем мире только 4 млн. получ али этот гормональны й препарат. О днако, у 5% воз никали аллергич еские реакции, об ус ловленны е антигенной несовместимос тью гормонаи клетокч еловека, т.е. такие б ольны е б ы - ли об реч ены нагиб ель. Киш еч ная палоч касо встроенны м геном инсулина синтез ируетв культуре до 200 г инс улинана1 литркультуральной среды , ч то эквивалентно колич еству инсулина, вы деленному из 1600 кг поджелу-
доч ной желез ы коровы или свиньи и не вы з ы вает аллергии. |
П риродны е |
ш таммы б актерий инс улинникогданепродуцировали. |
|
2. С О ЗДА Н И Е И С К РИ Н И Н Г Б А Н К А ГЕ Н О |
В |
Как уже отмеч алос ь, вы деление (получ ение) нужного гена(транс гена), намеч енного для перенос а– один из главны х этапов в генетич еской инженерии. Г енможетб ы ть вы делениз естественны х источ ников (из под-
19
ходя щ его генома), синтезированхимич еским путем (по имею щ ей с я последовательности нуклеотидов) или ф ерментативны м путем с использованием механиз маоб ратной транскрипции (синтез кД Н К наматрице мРН К с помощ ью об ратной транскриптаз ы ), получ ен с помощ ью полимераз ной цепной реакции (П ЦР). Ч ас тонужны й генвы деля ю тиз бан ка ген ов.
2.1. При нц и п ы создани я банка генов
Бан к ген ов, или клон от ека (ген ом н ая би бли от ека) – это коллекция клонов Д Н К, вклю ч аю щ ая все ф рагменты , входя щ ие в состав геномаданноговида.
Д ля ее соз дания необ ходимо вы деление всей (тотальной ) геномной Д Н К, ее ф рагментация с помощ ью рестриктаз или методом дроб овика (ультразвуком); присоединение получ енны х ф рагментов к клонирую щ им векторны м молекулам, введение рекомб инантны х Д Н К в реципиентны е б актерии для их последую щ его клонирования . В рез ультатеполуч аю тклоны с раз ны ми ф рагментами одной молекулы Д Н К . Н аб ор клонированны х ф рагментов геномаи назы вается бан ком ген ов, аточ нее, - э то произвольная (случ ай ная ) коллекция клонированны х ф рагментов Д Н К, представля ю щ ая соб ой с овокупность всех нуклеотидны х последовательностей Д Н К данного индивида или вида. О днажды получ енная , б иб лиотекагенов может храниться и использ оваться неогранич енно долго. В б иб лиотеке содержится вся нас ледственная инф ормация организ ма. Банк генов – это не только ис точ ник для получ ения нужного транс гена, но и источ ник материаладля из уч ения структуры , ф ункции и регуля ции индивидуальны х генов, с труктуры и ф ункции б елков. С его помощ ью можно также реш ить проб лему сохранения геноф ондаисч ез аю щ их видов.
Г ены эукариотз анимаю тдос таточ но протя женны е уч ас тки Д Н К (до 2,5 млн пн). Д ля клонирования таких крупны х ф рагментов плаз мидны е векторы не подходя т. В э том случ ае использ ую т клонирую щ ие векторы , соз данны е на ос нове б актериоф ага λ (воз можны й раз мер клонируемого ф рагментадо 23тпн), космиды (до 45 тпн) или же искусственны е хромосомы (от100 до>1000тпн).
П ервую геномную б иб лиотеку соз дали Т. М аниатис с сотрудниками в 1978 г. О ни ис пользовали Д Н К из геномаD. melanogaster, которую клонировали в клетках E. coli.
Аналогич ны е коллекции, получ енны е из индивидуальны х хромосом или их ч астей , наз ы ваю тся хр ом осом н ы м и би бли от еками .
Би бли от еки кДН К составля ю т копии Д Н К, комплементарны е РН К (рисунок7). П оскольку кД Н К получ аю тиз з релы х мРН К , прош едш их процес синг, они не с одержат интронов. Биб лиотека кД Н К отражает с пектр генной активности в клетках, из которы х онаб ы лавы делена. С оздание таких б иб лиотекполез но для сравнения генной активнос ти в клетках разны х тканей .
20
|
|
|
|
|
|
|
|
|
п о ли (А ) х во ст |
||||||
м РНК |
5' |
|
|
|
|
|
|
|
|
|
AAAAAA3' |
|
|||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
Обратная |
|
|
ТТТТТТ |
5' |
|
||||
|
|
|
|
|
|
|
|
|
п рай м ер |
||||||
|
|
|
|
|
|
транскри п ци я |
|
||||||||
|
|
|
|
|
|
|
о ли го |
(dT) |
|||||||
К о м п лекс |
5' |
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
AAAAAA3' |
|
||||||
м РНК :кДНК |
3' |
|
|
|
|
|
|
|
|
|
ТТТТТТ |
5' |
|
||
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
Разру шени е м РНК |
|
|
||||||||
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
РНК -азо й Н |
|
|
|
|
|
|
|
|
|
5' |
|
|
|
|
|
|
|
|
|
AA 3' |
5' |
|
||
|
3' |
|
|
|
|
|
|
|
|
|
ТТТТТТ |
|
|||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
До страи вани е вто ро й цеп и |
ДНК -п о ли м еразо й |
I |
||||||||||||
|
с и сп о льзо вани ем фрагм енто в РНК в качестве |
||||||||||||||
|
п рай м еро в |
|
|
|
|
|
|
3' |
|
||||||
|
|
|
|
|
|
|
|
||||||||
|
5' |
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
ТТТТТТAAAAAA5' |
|
||||
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
||||||
|
3' |
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
ДНК -ли газа сши вает |
|
|
|||||||
Дву х цеп о чечная |
|
|
|
|
|||||||||||
|
|
фрагм енты ДНК |
|
|
|||||||||||
кДНК |
5' |
|
|
|
3' |
|
|||||||||
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
5' |
|
||
|
3' |
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
||||
Рисунок7. С интез двухцепоч еч ной кД Н К намРН К |
|
||||||||||||||
У эукариот только мРН К содержит поли-А хвос ты . |
И спольз уя э то |
||||||||||||||
кач ес тво, мРН К вы деля ю т. Затем in vitro кэтой мРН К доб авля ю ткороткую цепь олиго (dT), которая пос леотжигаслужитпрай мером для дей с твия об - ратной транскриптаз ы , синтез ирую щ ей комплементарную цепь Д Н К на
молекуле мРН К. Д алее с помощ ью РН К -аз ы Н |
разруш аю т мРН К в ком- |
плексе мРН К :кД Н К. О стровки полуразруш енной |
мРН К с лужатпрай мера- |
ми для синтез авторой цепи Д Н К с помощ ью Д Н К-полимераз ы I по матри- |
|
це кД Н К. Ф рагменты новой Д Н К -цепи с ш иваю тся с помощ ью Д Н К - |
|
лигаз ы . П ос ле того какмолекулы кД Н К синтезированы , кним с помощ ью Д Н К -лигазы “приш иваю т”липкие концы , пос ле ч его встраиваю тв клонирую щ ий вектори перенос ятся для их размножения в б актерии.
Таким |
об раз ом, б иб лиотека генов представля ет соб ой наб ор ф раг- |
ментов Д Н К |
, встроенны х в вектор. |
2.2. В ыбор нуж ногогена и зклонот еки (скри ни нгбанка генов)
П оискнужны х генов в смес и клонированны х ф рагментов Д Н К б иб - лиотеки ос ущ ествля ется различ ны ми способ ами; суть всех их состоит в скринировании б иб лиотек. Рас смотрим некоторы еиз них.
1. Е сли нам доступен ис комы й ген, мы з наем его местоположение, молекуля рную масс у, то его можно вы делить путем разделения ф рагментов по молекуля рной масс е и з аря ду при помощ и гель-электроф орез а. Д ля