

Генетическая инженерия и безопасность
.pdf21
этого ис следуемы е об раз цы (с одержащ ие раз лич ны е ф рагменты одной и той жеД Н К ) нанос ятсверху наагарозны й или полиакриламидны й (П ААГ ) гель. П ри наложении нагель электрич ес кого поля ф рагменты нач нутперемещ аться вниз (ототрицательногокположительному полю су, пос кольку
молекулы Д Н К отрицательно заря жены ) со с коростью , |
з авися щ ей отдли- |
ны (массы ) ф рагмента. Э то с вя зано с тем, ч то в гелевой |
среде, состоя щ ей |
из пор, молекулы Д Н К раз ного раз мератратя тразное время напреодоление пор. Ч ем меньш е раз мерф рагментов, тем б ы стрееони движутся . В результате электроф орез ав геле об раз уется ря д полос, рас положенны х одна под другой . В ерхние полосы соответствую тф рагментам, имею щ им б олее
крупны е раз меры , а нижние – ф рагментам с |
б олее мелкими раз мерами. |
П олосы вы я вля ю тся при окраш ивании гелей |
б ромис ты м э тидием и про- |
смотре гелей в ультраф иолетовом свете (рисунок8). Ч тоб ы определить относ ительную молекуля рную масс у раз деленны х ф рагментов, одновременно проводя т электроф орез маркерны х молекул с из вестны ми молекуля р- ны ми мас сами. Зная , какую мас су имеет ф рагмент, содержащ ий интересую щ ий нас ген, можно вы делить его из электроф оретич еского геля и ис - пользовать поназнач ению .
2. Е сли единственны м “паспортом”искомого гена с лужит его нуклеотидная последовательность, то поискнужного генав смес и ф рагментов Д Н К осущ ествляю тс помощ ью методагиб ридиз ации нуклеиновы х кислот (такназ ы ваемой in situ – гиб ридиз ации). Д ля этого применя ю тм олекуляр - н ы езон ды . Зон ды – это искусс твенно синтез ированны е меч ены е (из отопа-
ми или ф луоресцентны ми крас ителя ми – |
химич ес ки) неб ольш ие (10-30 |
нуклеотидов) сегменты одноцепоч еч ной |
Д Н К (или РН К, или ее Д Н К - |
копии), комплементарны е ис комому гену. Зонд – это с интетич еский олигонуклеотид (короткий с егментодноцепоч еч ной Д Н К) с из вестной нуклеотидной последовательностью , которы й использ уется для вы я вления комплементарны х пос ледовательностей с помощ ью гиб ридизации (т.е. зонды
служатиндикаторами гомологии при гиб ридиз ации с оответствую щ |
их по- |
следовательнос тей ). Д ля успехараб отпо генетич еской инженерии |
важно, |
ч тоб ы кажды й |
конкретны й зонд представля лкопии одной молекулы Д Н К |
с известной |
последовательностью нуклеотидов, а данны е по Д Н К - |
гиб ридиз ации во всех лабор ат ор и ях м и р а можно б ы ло сравнивать между соб ой .
Реакция гиб ридиз ации нуклеиновы х кислот– ч увствительны й метод вы я вления с пециф ич еских пос ледовательностей нуклеотидов. Г иб ридиза-
ция in situ – этоотжиг одноцепоч еч |
ногоф рагментаД Н К накомплементар- |
ны й ему уч ас ток другой молекулы |
Д Н К с об раз ованием двухцепоч еч ной |
гиб ридной молекулы . О тжиг – процесс восстановления (ренатурации) двухцепоч еч ны х молекул Д Н К из одиноч ны х полинуклеотидны х цепей путем постепенногоохлаждения .
П роцедура поиска нужны х генов в б анке получ ила наз вание блот - т и н га (отангл. blotting - промокание). Блот т и н г – это метод перенесения электроф оретич еских ф рагментов Д Н К наспециальную пленку (мемб рану)
22
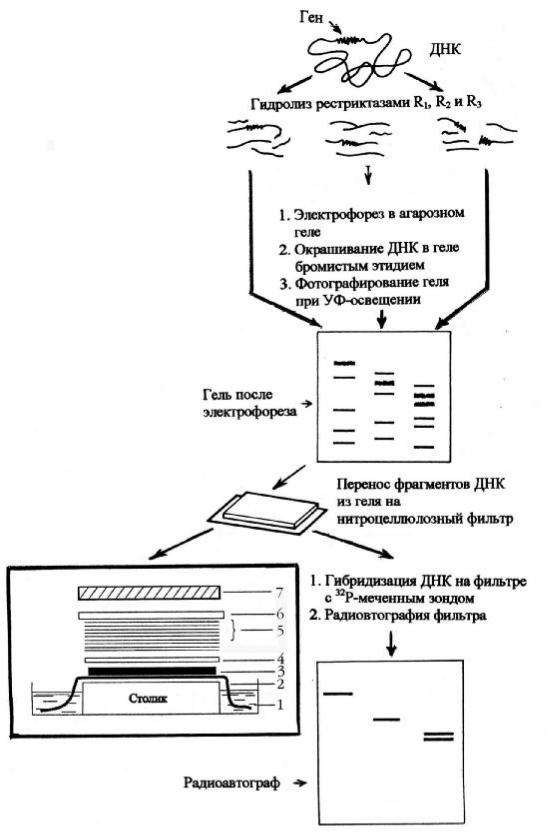
Рисунок8. С хемаб лоттингапоС ауз ерну:
1 – б уф ер; 2 – ватман; 3 – гель; 4 – нитроцеллю лозны й ф ильтр; 5 и 6 – ф ильтровальная б умага; 7 – стекло; 8 – груз (0,5 кг)
23
из нитроцеллю лоз ы , свя з ы ваю щ ую (иммоб илиз ую щ ую ) одноцепоч еч ны е молекулы Д Н К .
Саузер н -блот т и н г (по ф амилии предложивш его его автора) основан наперемещ ении ф рагментов Д Н К б лагодаря капилля рному эф ф екту. П роцес с переносаф рагментов Д Н К, находящ ихся в агароз ном геле, напленку из нитроцеллю лозы с помощ ью ф ильтровальной б умаги похож напромокание.
Анализ проводя тс ледую щ им об разом (рисунок8).
- В ы деленную , оч ищ енную , денатурированную и раз б итую наф рагменты Д Н К помещ аю т на лист агароз ного геля, где происходитэ лектро-
форетич ескоераз делениеф рагментов помас сеи заря ду.
-Л ист агароз ного геля , где произош ло электроф оретич еское ф рак-
ционирование смеси ф рагментов Д Н К |
по мас се и заряду, |
помещ аю т на |
ф ильтровальную б умагу, смоч енную |
концентрированны м |
с олевы м (б у- |
ферны м) рас твором.
-Затем нагель наклады ваю тнитроцеллю лозны й ф ильтр, где проис -
ходит иммоб илизация (или адсорб ция , или ф икс ация ) одноцепоч еч ны х ф рагментов Д Н К .
- П оверх ф ильтранаклады ваю тстопку лис тов сухой ф ильтровальной б умаги, которая об ес печ ивает медленны й ток б уф ерного раствора ч ерез гель (т.е. служит своеоб раз ны м капилля рны м нас осом). С олевой раствор, проходя ч ерез агарозны й гель, увлекаетз ас об ой ф рагменты Д Н К , которы е задерживаю тся нитроцеллю лозой , и свя зы ваю тся с ней , арас творвпиты вается сухой ф ильтровальной б умагой .
- Д алее Д Н К денатурирую тщ елоч ью , а ф ильтр вы держиваю тв вакууме при температуре 800С , в рез ультате ч его одноцепоч еч ны е ф рагменты Д Н К необ ратимо иммоб илиз ую тс я (ф иксирую тся ) нанитроцеллю лоз е. П ри этом расположение полос иммоб илиз ованной Д Н К точ но соответствуетих расположению в геле.
- Д Н К, свя занную с ф ильтром, помещ аю тв рас творс меч ены м Д Н К зондом, в котором и проис ходитгиб ридиз ация . Г иб ридиз ироваться (об разовы вать водородны е свя з и) со специф ич еским зондом б удуттолько комплементарны е ему ф рагменты Д Н К, которы е можно об наружить в виде светлы х полос на рентгеновс кой пленке, т.е. радиоавтограф ии нитроцеллю лоз ногоф ильтра(рисунок8).
Д ля вы деления и анализ аРН К |
(например, для вы я снения того, при- |
сутствуетли в данном типе клеток |
мРН К, с ч итанны е с данного гена, т.е. |
экспресс ируетс я ген или нет; для определения колич ес тваэтой |
РН К и его |
|
из менения в развитии |
данного типа клеток; для определения размера |
|
транскрипта какого-то |
гена и др.) применя етс я Нозер н -блот |
ан али з, во |
многом похожий |
наСаузер н -блот т и н г. В данном с луч ае молекулы РН К, |
вы деленны е из |
клетки, раз деля ю тся по размерам с помощ ью гель- |
электроф орез а, аз атем перенос ятся наф ильтр. П ос ле гиб ридиз ации с ме- ч ены м одноцепоч еч ны м з ондом вы я вля ю тс я местагиб ридиз ации (гомологии) РН К и зонда.
24
3. Е с ли нуклеотидная последовательность искомого гена(или мРН К ) не из вес тна, но из вестен б елок, синтез которого он контролирует, то то можно вы делить неб ольш ое колич ество ч ис того б елка, определить амино-
кис лотную последовательность некоторой |
его ч асти (достаточ но знание 5- |
6 аминокислотны х остатков). П ользуя сь |
таб лицей генетич еского кода, |
можно установить все воз можны е последовательнос ти нуклеотидов в том уч астке мРН К (или с амого гена), которы й кодируетданную аминокис лотную последовательнос ть. В этом случ аеможно синтез ировать з онд для поисканужны х клонов в б иб лиотекегенов.
С ущ ествую ти другие способ ы вы деления нужны х генов из клоноте-
ки.
3. Г Е Н Н А Я И Н Ж Е Н Е РИ Я М И К РО О РГА Н И ЗМ О В
М етоды генной инженерии наиб олее детально разраб отаны намикроорганиз мах. Н ач алом промы ш ленной генной инженерии микроорганиз - мов приня то сч итать 1980 г, когда в С Ш А б ы л вы дан первы й патент на генно-инженерны й ш тамм микроорганиз ма, с пособ ны й разлагать неф ть.
Е щ еч ерез 2 годаб ы лраз реш ендля клинич еского использования |
получ ен- |
ны й из б актерии первы й лекарственны й препарат– ч еловеч еский |
инс улин. |
Более 20 ф ирм Я понии и нес колько Американс ких ф ирм разраб отали другой оч ень важны й медицинский препарат– интерф ерон, которы й эф ф ективен при раз лич ны х вирусны х з аб олевания х и з локач ественны х новооб разованиях. В наш ей стране Ю .А. О вч инников и В .Г . Д еб аб ов с сотрудниками получ или микроорганиз мы , эф ф ективно синтез ирую щ ие интерф ерон ч еловека(до 5 мг интерф ерона на1л суспенз ий б актерий , ч то в 5000 раз б ольш е, ч ем с одержится в 1 литре крови доноров). В С Ш А около 63% медицинских препаратов производится с помощ ью б иотехнологич еских методов, в с транах Западной Е вропы – 25%, в Я понии – 7%.
П римером генной инженерии я вля етс я также получ ение б актерии Е. соli со встроенны м геном соматотропина– гормонаростач еловека, которы й ис польз уется не только в медицинс ких целях, но и в практич еском животноводстве, повы ш ая с егопомощ ью интенс ивность ростаживотны х.
В современной б иотехнологии ш ироко ис пользую тс я трансгенны е микроорганиз мы , продуцирую щ ие лекарс твенны е препараты : антиб иотики, гормоны , ф ерменты , витамины ; вакцины против инф екционны х з аб о- леваний ; раз лич ны е диагностич еские препараты для диагностики нас ледственны х и инф екционны х б олезней (например, В И Ч , вирусного гепатита и др); для производстванез аменимы х аминокислот, б иодоб авоки др.
4. ГЕ Н Н А Я И Н Ж Е Н Е РИ Я РА С ТЕ Н И Й
4.1. Агробакт ери альная т рансф орм ац и я
П римеры об разования транс генны х растений в природны х ус ловиях ш ирокоизвестны .
25
О пухолевоез аб олевание, из вестноекаккоронч аты й галл, описалещ е Аристотель. В 1907 г. Э . Cмити К. Таунс енд показали, ч тоэтозаб олевание вы зы вает поч венная б актерия Agrobacterium tumefaciens. В ы деленная в ч ис той культуре, этаб актерия можетприводить к об разованию опухолей (какправило, у корневой ш ей ки) у многих голосеменны х и покры тосеменны х растений , ч то по сущ ес тву можетрасс матриватьс я какприродная ген- но-инженерная с истема(рисунок9).
В 70-х годах Д ж. Ш елли др. вы явили, ч то прич иной опухолеоб разования я вляю тся такназы ваемы е Ti-плазмиды (отангл. tumor inducingин- дуцирую щ ая опухоль), об наруженны е в клетках некоторы х ш таммов A. tumefaciens. Ti-плазмидапроникаетиз клетки б актерии в растение и ч асть ее, назы ваемая Т -Д Н К (от англ. transferred DNAперенос я щ ая ся ), ковалентно встраивается в хромосомы инф ицируемого растения . В природе этотф рагментпереноситгены , которы е с пособ ствую тразмножению агро- б актерий и даю тим возможность параз итировать напораженном растении.
Г ены , входя щ ие в сос тав T-Д Н К , ф ункционирую тлиш ь после их пе-
реносав растительную клетку. Будуч и интегрированной |
с хромосомой , Т - |
Д Н К индуцируетв месте з аражения неконтролируемы й |
рос тнедиф ф ерен- |
цированны х клеток, вы з ы вая об разование опухоли (коронч аты х галлов, напоминаю щ их раковы е клетки животны х), гиперпродукцию ф итогормонов: цитокининов и индолилуксус ной кис лоты - И У К (аукс ина), а также синтез рядапроизводны х аминокислот, об ъ единенны х под об щ им термином оп и н ы , которы х нетв з доровы х клетках ни у одногорастения .
При культивировании в условиях in vitro клетки опухоли могутрас ти
вотс утс твие с пециальны х гормонов (ауксинов/цитокининов), необ ходи-
мы х |
для культивирования нормальны х растительны х клеток, |
поскольку |
|
эти |
гормоны клетки |
опухоли с интез ирую т сами. О пухоль |
воз никает |
вс ледствие наруш ения |
б аланса ф итогормонов, от которого з ависит нор- |
||
мальны й морф огенез растения . Т.е. эторезультатф ункционирования онкогенов, продуктами которы х явля ю тся ф итогормоны (ауксины и цитокинины ). О пины , вы деля емы еклетками опухоли, б актерия использ уетв кач естве источ ников углеродаи азотадля с воегорос таи размножения . С амаб актерия в клетку не проникает, аос тается в межклеточ ном прос транстве и использ ует клетки со вс троенной T-Д Н К как ф аб рику, продуцирую щ ую опины . Ti-плаз мидаотнос ится ккласс у конъ ю гативны х плаз мид.
Д оказ ательством того, ч то именно Ti-плазмиды , ане гены хромосомы б актерии ответственны з аподдержание транс ф ормированного сос тоя - ния клетоккоронч аты х галлов, я вля ется то, ч то если б ы агроб актерии содержали мутантны е Ti-плазмиды , не происходило б ы ни з аражения , ни об - разования коронч аты х галлов, ни синтезаопинов.
Ti-плазмиды расс матриваю тс я как природны е векторы , поскольку могутпередаваться отб актерии в клетки растений в природны х ус ловия х. Т.е. вз аимоотнош ения б актерий с рас тения ми представляю т особ ы й вид параз итиз ма, когдаб актерия не просто использ уетпитательны е вещ ества растения -хозя ина, азаставля етрастительны е клетки из менить свой мета-
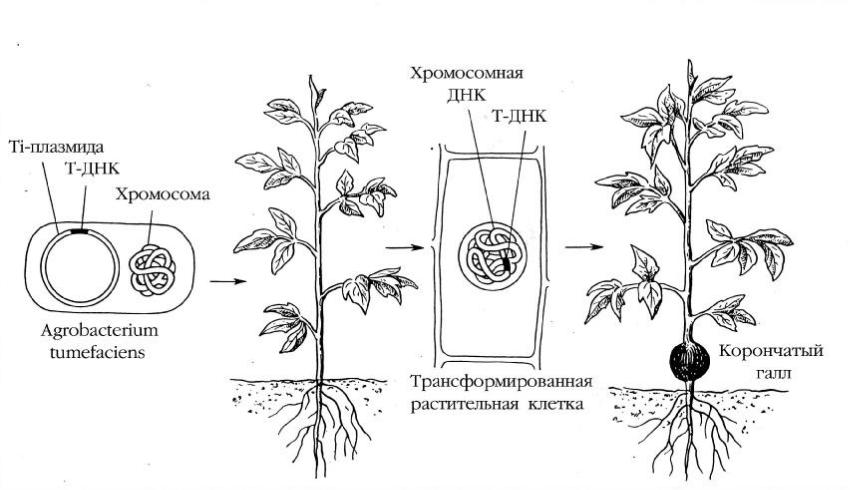
Рисунок9. “Г енетич еская колонизация ”вы сш егорастения б актерией A. tumefaciens (П ируз я н, 1988).
A. tumefaciens сущ ествуетв ризосф ерерастения . В клетках б актерии наряду с хромосомой содержится Ti – плазмида, которая проникаетв клетку растения и ч асть ее, T-Д Н К, встраиваетс я в геном растения, приводя коб разованию опухоли и синтез у опинов
27
б олизм и с интез ировать те вещ ества(опины ), которы е необ ходимы агро- б актерия м. Такие отнош ения A. tumefaciens и растения Ш елл назвал генетич еской колониз ацией , которая представля етсоб ой экспериментпо генной инженерии, поставленны й самой природой .
Ti-плазмидаоказалась идеальны м природны м вектором для введения ч ужеродны х генов в клетки растения . Н аееосновев условия х эксперимента с оздаю тс я искусственны е векторы . Д ля этого генетич ескую конструкцию , содержащ ую ген, намеч енны й для переноса, встраиваю т в T-Д Н К (рисунок10).
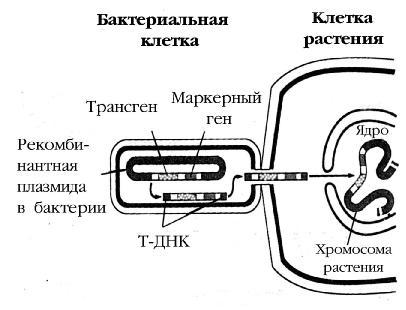
Рис унок10. С хемаагроб актериальной трансф ормации растительной клетки [Alberts,1994]:
О б ласть T-Д Н К, содержащ ая трансген(это, например, можетб ы ть ген устой ч и- вости кгерб ицидам, или вредны м насекомы м и др.), маркерны й ген(например, ген устой ч ивости к антиб иотику канамицину) и все необ ходимы е регуля торны е последовательности и сай ты рестрикции, вы резается из рекомб инантной Ti-плазмиды агроб актерии, переносится в растительную клетку, гдевс траивается в хромосомурастения .
Ч топредставля етсоб ой об лас ть T-Д Н К?
Раз мер всей Ti-плаз миды с оставля ет 200-250 тпн, раз мер T-Д Н К в раз ны х плаз мидах варьирует от 10 до 30 тпн (ч то составля ет примерно 10% Ti-плаз миды ). Н аT-об ласти картировано не менее 7 генов, кажды й из которы х регулируетс я соб ственны м промотором (ч то сходно с генами эукариот). Э ти гены отвеч аю тз ас интез опинов (тип которы х, например, нопалин, октопин, агроцинопин, манопин з ависитот ш тамма агроб актерии, вы зы ваю щ его его об разование) и подавление диф ф еренцировки клеток (онкогены ) – подавление об раз ования корней и поб егов. Э ти гены ф ункционирую т лиш ь после их переноса в растительную клетку. С концов T- Д Н К огранич енаправы м и левы м пря мы ми повторами из 25 пн, ч топридаетей сходствос моб ильны ми генетич ескими элементами (рис унок11). П о-
28
этому лю б ая Д Н К, вставленная между этими повторами, б удетприня таз а T-Д Н К и перенес енав растительную клетку. В ажноотметить, ч то все гены (их около 35), ответственны е заперенос и интеграцию T-Д Н К , находя тся нев T-Д Н К , ав об ласти вирулентности (vir-об лас ть), рисунок11.
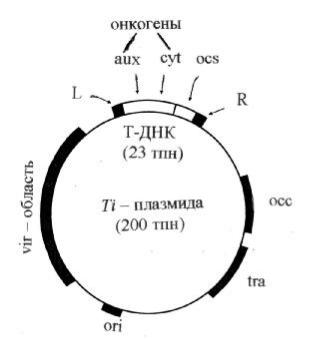
Рисунок11. Г енетич еская картаTi-плазмиды октопиновоготипа
T-Д Н К (трансф ормирую щ ая Д Н К) содержитгены ауксина(aux), цитокинина(cyt) и опина(ocs), которы е транскриб ирую тс я и транслирую тс я только в растительны х клетках; vir-об ласть, содержащ ая гены , продукты которы х об еспе- ч иваю твы резаниеи перенос Т-Д Н К в растительную клетку; tra-об ласть, где локализованы гены , контролирую щ ие конъю гацию б актерий ; ori – сай тинициации репликации, об еспеч иваю щ ий репликацию и стаб ильное поддержаниеплазмиды в A. tumefaciens; occ – гены , кодирую щ ие ф ерменты катаб олизмаопина; L и R – левая и правая ф ланкирую щ иепоследовательности T-Д Н К соответственно.
О днако практич еское ис пользование природны х Ti-плаз мид каквекторов для переноса генетич еской инф ормации в растительны е клетки и клонирования генов затруднено из -з аееб ольш их раз меров (до250 тпн, тогдакак для прокариотвектор pBR322 имеетраз меры вс его 4,4 тпн). У ч е- ны ми б ы ли разраб отаны раз лич ны е стратегии введения ч ужеродны х генов в состав T-Д Н К , однаиз которы х, наш едш ая ш ирокое применение, – соз - дание б инарной векторной с истемы . В этом с луч ае конструирую тдвавектора, совмес тно взаимодей ствую щ ие друг с другом, один из которы х содержитоб ласть T-Д Н К , адругой – гены vir-об лас ти, об еспеч иваю щ ие вс е ф ункции переносаT-Д Н К в геном растительны х клеток. Трансгены , кодирую щ ие хоз я й ственно ценны е приз наки, встраиваю т в T-Д Н К (рисунок 11). Э таже об лас ть с наб жается маркерны ми генами (для отб оратрансф ормированны х рас тительны х клеток), э укариотич еским промотором (уз наваемы м растительны ми полимераз ами, например35S-промоторвирусамо- заики цветной капусты - CAMV) и уникальны ми с ай тами рестрикции (в ко-
29
торы е встраиваю тс я ч ужеродны е ф рагменты Д Н К ). П рич ем, гены , подавля ю щ ие диф ф еренцировку растительны х клеток и вы зы ваю щ ие развитие опухоли (онкогены ), инактивирую тся или вы рез аю тс я, вследствие ч его рос ттранс ф ормированны х растений не наруш аетс я . Таким об разом, плаз - мидаи агроб актерия нач али раб отать наполуч ениетранс генны х растений .
В последние годы для соз дания искус ственны х векторов ис пользуется Ri-плаз мида(отангл. root inducing – индуцирую щ ая корни), присутствую щ ая в ш таммах Agrobacterium rhizogenes. Ri-плаз миды вы годно отли- ч аю тс я отTi-плаз мид тем, ч то они я вля ю тс я естественны ми б езвредны ми векторами, т.е. после встраивания T-Д Н К в хромосомную Д Н К растительны х клеток в об ласти з аражения наб лю даетс я усиленное об разование кореш ков (“б ородатость”), из которы х легч е регенерировать з доровы е плодовиты ерастения , ч ем из недиф ф еренцированной ткани опухоли.
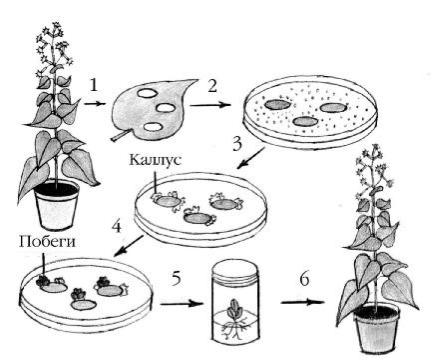
Как напрактике осущ ес твля ю тгенетич ескую транс ф ормацию рас - тительны х клеток? Н еоб ходимы м ус ловием для инф екции Ti-плаз миды я в- ля етс я поранение растения . П оэ тому агроб актерии, содержащ ие рекомб и- нантны е плазмиды , нанося т на срезанную ч асть растения (например, по- б ег) или ос ущ ествля ю тсовместное культивирование (кокультивирование) агроб актерий и с терильного растительного материала (например, сегментов междоуз лий , листовы х дисков, протоплас тов) напитательны х средах в ус ловия х in vitro. Н а рисунке 12 приведен типич ны й пример получ ения
транс генного растения |
таб ака путем агроб актериальной транс ф ормации |
лис товы х дисков. |
|
О днако, несмотря |
на эф ф ективность агроб актериальной трансф ор- |
мации, круг хоз я ев агроб актерий огранич ен. Агроб актерии об ладаю тспособ ностью интегрировать свой генетич ес кий материалпреимущ ественно в клетки двудольны х растений . Альтернативны ми с пособ ами преодоления проб лемы огранич енного круга растений , ч увствительны х к трасф ормации, я вля ю тс я методы пря мого переносач ужеродной Д Н К - микроинъ екция , электропорация , б омб ардировкамикропуля ми (один из с амы х эф ф ективны х методов для трансф ормации однодольны х растений ) и др.
4.2. В ект оры на основе хлороп ласт ной и м и т охондри альной ДН К
Н е менее перс пективны м для соз дания векторов я вля ется ис пользо-
вание хлороплас тной |
(хп) и митохондриальной (мт) Д Н К (т.е. внея дерной , |
цитоплазматич еской |
Д Н К ). Э то свя з ано с тем, ч то растительная клетка |
можетсодержать б ольш ое колич ество копий (до 50 ты с .) хп Д Н К или мт Д Н К. П оэ тому перенос ч ужеродны х генов в их составе приведеткнакоплению б ольш ого колич естваб елкового продуктапо сравнению с ф ункционированием этого же генав сос таве я дерной Д Н К. Кроме того, для б оль- ш инс твавидов растений с ущ ествуетматеринское нас ледование цитоплаз - матич еских генов. П оэтому использование генетич еских конс трукций на основе хпД Н К или мтД Н К исклю ч аетвоз можность прис утствия в пы льце ч ужеродны х генов и огранич иваетих неконтролируемоераспространение
30
Рис унок12. Э тапы получ ения транс генны х растений таб акапутем агроб актериальной трансф ормации лис товы х дис ков [Alberts,1994]:
1 – из листатаб акавы резаю тдиски, которы е помещ аю тв ч аш ки П етри со специальной питательной средой ; 2 – листовы е диски инкуб ирую тс агроб актериями, содержащ ими рекомб инантны е плазмиды с трансгеном; 3 – отб ортранс - ф ормантов (вклю ч ивш их в свой геном трансген) населективной среде и индукция каллус а(ткани, состоящ ей из активно делящ ихс я недиф ф еренцированны х = дедиф ф еренцированны х клеток); 4 – 5 – регенерация целогорастения из каллус а листовы х дисков: при перенос е каллусной ткани наморф огенную среду, проис - ходитоб разование поб егов (4), апри переносе последних насреду для укоренения развиваю тс я корни (5); 6 – проб ироч ное растение с корнями перенося т в поч ву, оносодержиттрансген, придаю щ ий растению новы й признак.
(например, генов ус той ч ивос ти к герб ицидам от культурны х растений к сорня кам с помощ ью пы льцы ). Т.е. растения , получ енны ес их ис пользованием, б олее б езопасны для окружаю щ ей с реды по сравнению с об ы ч ны ми транс генны ми растения ми.
Рас смотрим преимущ ества и особ енности с оздания векторов наос - новехпД Н К.
П ластиды находя тся в б ольш ом колич естве в разны х органах и тканя х растений . Г еном плас тид (п ласт ом ) – кольцевая молекула двухцепо- ч еч ной Д Н К раз мером 120-180 тпн. В каждой пластиде содержится от10 до 100 пластом. Е динич ная клеткалистаможетсодержать до 100 пластид, ас ледовательно, до10 ты с плас тидны х геномов. В состав кольцевы х молекул входя тгены рРН К и тРН К, атакже гены , продукты которы х необ ходимы для ф ункционирования хлоропластов. Репликация и транскрипция хлоропластного генома осущ ествляетс я автономно от я дерного, ч то дает воз можность использовать хпД Н К в кач ествевектора.