

л а нванов и мам одев
РЕСНИЧНЫЕ ЧЕРВИ
ИХ ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИ
Я
АКАДЕМИЯ НАУК СССР ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
А. В. ИВАНОВ и Ю. В. МАМКАЕВ
РЕСНИЧНЫЕ ЧЕРВИ (TURBELLARIA)
ИХ ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ
ФИЛОГЕНЕТИЧЕСКИЕ ОЧЕРКИ
ИЗДАТЕЛЬСТВО «НАУКА» Ленинградское отделение Ленинград • 197
3Ресничные черви (Turbellaria), их происхождение и эволюция. Филогенетические очерки. Иванов А. В. и Мамка ев Ю. В. 1973. Изд-во «Наука», Ленингр. отд., Л. 1—221.
На основе сравнительно-морфологического анализа класса ресничных червей дается методологически обоснованное эволюционное объяснение их организации и онтогенеза. Однако основная цель книги — обсуждение проблем филогенетической зоологии, связанных с выяснением происхождения и систематического положения этой группы животных. От толкования этих проблем зависит решение многих рассмотренных в книге вопросов современной эволюционной морфологии, в первую очередь — выяснение природы первичной гистологической структуры и начальных этапов эволюции покровов, кишечника, нервного, выделительного и полового аппаратов, а также спирального дробления и мезодермы. Авторы развивают планулоидную теорию происхождения Turbellaria в том ее варианте, который был намечен еще И. И. Мечниковым. Критически анализируются теории регрессивной эволюции плоских червей. Приводятся оригинальные схемы прототипа Turbellaria и филогенетических связей между отрядами этого класса. Илл. — 115, табл. — 1, библ. — 391 назв.
Ответственный ре да к т о р А. А. СТРЕЛКОВ
Й
5^q2^73
781~73 © Издательство «Наука», 197
3Посвящается светлой памяти Владимира Николаевича Беклемишева
ПРЕДИСЛОВИЕ
В предлагаемых читателю очерках обсуждаются основные и спорные вопросы системы и филогении ресничных червей — Turbellaria. Для этого требуется, разумеется, подробное рассмотрение современных данных по сравнительной анатомии и эмбриологии этих животных. Наша работа не претендует на всестороннее и полное освещение класса ресничных червей. Это не сводка, но морфологическая монография; материалы по экспериментальной зоологии и вопросы экологии, зоогеографии и систематики низших таксономических групп нами не рассматриваются.
Ресничные черви, по убеждению многих зоологов, представляют собой одну из важнейших в эволюционном отношении примитивных групп многоклеточных животных, хотя эту истину, как ни странно, ныне приходится усиленно защищать. Выяснение происхождения и изучение филогении турбеллярий тесно связаны с решением многих существенных проблем эволюции низших Metazoa.
Особенно много споров вызывают бескишечные турбеллярии — Acoela. Одни зоологи, особенно те, кто изучал Acoela, рассматривают их как наиболее примитивный отряд ресничных червей и, следовательно, как самую примитивную группу Bilateria. Другие пытаются доказать вторичную упрощенность их организации.
Более 15 лет тому назад из печати вышла известная книга
Н. А. Ливанова «Пути эволюции животного мира» (1955), в которой автор отстаивал неприемлемую, с нашей точки зрения, концепцию о вторичной деградации Acoela и производил турбеллярий от гастрееподобных предков. В 1961 г. известный немецкий турбелляриолог, проф. Петер Акс (Р. Ах), ученик проф. А. Ремане (A. Remane), опубликовал большую статью об эволюции турбеллярий, в которой осветил исходя из господствующей в Западной Европе теории Ремане о вторичном упрощении плоских червей, произошедших якобы от метамерных целомических предков, вопросы, связанные с происхождением ресничных червей. В 1963 г. Петер Акс опубликовал эту стат№ Я на английском языке в США в книге «The lower Metazoa» (издание Калифорнийского университета), после чего она стала основным и наиболее авторитетным сочинением по вопросам происхождения и эволюции турбеллярий. Мы, однако, придерживаемся других взглядов и рассматриваем организацию турбеллярий с иных точек зрения. Это дает нам основание предлагать вниманию читателя данную работу.
Наши собственные представления об эволюции низших Metazoa формировались под влиянием идей И. И. Мечникова главным образом в процессе работы над лекциями по сравнительной анатомии беспозвоночных, читавшимися нами в течение многих лет для студентов-зоологов Ленинградского государственного университета. Вопрос о происхождении многоклеточных животных был рассмотрен в специальной книге (Иванов, 1968) именно с позиций мечниковской теории Фагоцителлы, которая имеет первостепенное значение для понимания природы и эволюции ресничных червей; В основу наших исследований по морфологии низших турбеллярий Acoela, начатых уже довольно давно (Иванов, 1952а; Мамкаев, 1965, 1967), положено учение И. И. Мечникова о фагоцитобласте и основанные на нем представления В. Н. Беклемишева о первичной ткани — смешанной паренхиме и о закономерностях ранней эволюции кишечника.
Все теоретические вопросы, затронутые в предлагаемых читателю очерках, мы обсуждали совместно, стараясь найти согласованное решение даже в тех случаях, когда наши точки зрения в той или иной мере расходились. Надеемся, что в большинстве случаев нам это удалось. Ю. В. Мамкаев написал раздел о половом аппарате, остальные главы принадлежат А. В. Иванову.
Неоднократно мы пользовались ценными советами наших друзей и товарищей — академика Б. Е. Быховского, проф.
А. А. Стрелкова, кандидатов биологических наук Л. А. Евдонина, Ю. С. Миничева и Л. Я. Кулинича, за что приносим им сердечную благодарность.
Ежегодно появляются превосходные работы по морфологии и систематике ресничных червей (Turbellaria). Тем не менее эти животные до сих пор остаются недостаточно исследованными, и их углубленное изучение все еще составляет одну из неотложных задач современной зоологии. Действительно, от того или иного толкования организации ресничных червей в сильной степени зависит решение многих важных проблем филогении и кдассифи- кации низших Metazoa. Что это так, видно из простого перечисления теорий и гипотез, в которых турбеллярии находятся в центре внимания.
Большинство зоологов думает, что если не все Metazoa, то по крайней мере Eumetazoa имеют единое происхождение. Однако некоторые современные авторы (Pantin, 1951, 1960; Boyden, 1953; Greenberg, 1959; Nursall, 1959, 1962; Kerkut, 1960) придерживаются гипотезы полифилетического происхождения Metazoa, полагая, что все основные типы многоклеточных животных возникли независимо друг от друга от различных групп протистов или даже от различных эобионтов, т. е. доклеточных существ. С этой точки зрения, разумеется, между плоскими червями и другими типами лежит глубокая пропасть, и все сходства между ними должны считаться не более чем поверхностными аналогиями.1Недавно эту идею поддержал также Гейдельберг(Hendelberg, 1967) на основании изучения спермиогенеза у Plathelminthes. О совершенно изолированном положении плоских червей, по его мнению, свидетельствует организация их очень своеобразных двужгутиковых сперматозоидов.
Трактовка организации турбеллярий играет важную роль и в попытках обосновать теорию происхождения Metazoa. Так, по гипотезе целлюляризации, защищаемой в настоящее время преимущественно Хаджи, Штейнбеком и Гансоном (Hadzi, 1944, 1963; Steinbock, 1952, 1958, 1963; Hanson, 1958, 1963), самые примитивные Eumetazoa — бескишечные ресничные черви (отрядAcoela) — произошли от многоядерных инфузорий (Ciliata) посредством обособления участков цитоплазмы вокруг отдельных ядер. Эта концепция влечет за собой ряд далеко идущих общих выводов. Ее сторонники не согласны с основными положениями клеточной теории и теории зародышевых листков, с учением об эволюции индивидуальности в животном мире и с многими хорошо обоснованными выводами цитологии, протозоологии и сравнительной анатомии низших Metazoa. В частности, гипотеза целлю- ляризации предполагает плазмодиальное состояние тканей у Acoela, причем рассматривает эту особенность как первичную. Именно поэтому сравнительное изучение тканевой структуры у низших турбеллярий приобретает особую актуальность.
С другой стороны, и одна из гипотез колониального происхождения Metazoa, а именно гипотеза Фагоцителлы И. И. Мечникова, или, как ее иногда называют, планулоидная теория (Hyman, 1942, 1951; Hand, 1963), тоже придает большое значение филогенетической стадии «ацельного метазоона». Действительно, современная зоология вооружает нас все возрастающим количеством аргументов в пользу происхождения всех Metazoa от общего фагоцителлообразного прародителя, который в свою очередь произошел от колониальных зоофлагеллят из отряда Protomona- dida вроде известной Proterospongia haeckeli. От этого общего предка возникли, с одной стороны, гастрееподобные Cnidaria и Ctenophora, а с другой — билатерально симметричные бески- шечные турбеллярии (см.: Иванов, 1968).
С этой точки зрения и вопреки мнению защитников гипотезы целлюляризации первичные турбеллярии имеют не плазмодиальное, а клеточное строение; каждая их клетка отвечает целому, моноэнергидному простейшему; их половой процесс не может быть выведен из конъюгации инфузорий, но сходен с широко распространенной у Protozoa копуляцией; их онтогенез, в том числе дробление яйца, есть наследие протозойных колониальных предков. Бескишечные турбеллярии, наконец, отличаются первичным отсутствием эпителизованного кишечника, так как у них сохранился еще настоящий фагоцитобласт (Мечников, 1886), т. е. имеется пищеварительная паренхима, пищеварение в которой носит еще в основном внутриклеточный характер — осуществляется нередко в блуждающих клетках — фагоцитах (Graff, 1904—1908; Беклемишев, 1944; Иванов, 1952а).
Ряд важнейших проблем филогенетической и систематической зоологии, в частности вопросы о происхождении Bilateria, о природе сходств и различий между Protostomia и Deuterostomia,
о возникновении и эволюции спирального дробления, целома и сегментации, сводится по существу к проблеме происхождения ресничных червей.
Дальнейшее углубленное изучение Turbellaria и тщательный анализ путей их эволюции необходимы не только для выработки правильного взгляда на их происхождение, но и для выясценщродственный отношений внутри этого класса. До сих пор по этому важному вопросу нет единого мнения. За наиболее примитивную группу принимались самые различные отряды, а именно Polycladida (Lang, 1881, 1884; Wilhelmi, 1913), Acoela (Graff, 1882, 1904; Мечников, 1886; Bresslau, 1928—1933; Bresslau u. Reisinger, 1928—1933; Беклемишев, 1937, 1944, 1963, 1964; Meix- ner, 1938; Westblad, 1948; Hyman, 1951; Иванов, 1952a, 1968; Мамкаев, 1967, 1968), Catenulida (Karling, 1940) и гипотетические формы, близкие к Macrostomida (Ах, 1961, 1963а).
Соответственно предлагались сильно различающиеся схемы филогенетических связей внутри класса и разные классификации Turbellaria.
В настоящих очерках мы хотим рассмотреть особенности организации и развития в разных отрядах турбеллярий, чтобы получить достаточно обоснованные выводы о их происхождении, эволюции и положении в системе животного мира. Как уже упоминалось, Петер Акс опубликовал блестящие статьи (Ах, 1961, 1963а), в которых он, однако, выступил как убежденный сторонник теории Spiralia, считающей ресничных червей сильно упрощенными потомками сегментированных целомических животных.
По многим проблемам, связанным с эволюцией турбеллярий, неоднократно высказывался большой знаток этих животных — В. Н. Беклемишев. Его посмертная статья (19636) в книге «The lower Metazoa» особенно интересна и имеет непосредственное отношение к нашей теме. Среди других значительных работ русских зоологов, в которых анализируется организация ресничных червей, следует отметить книгу Н. А. Ливанова (1955) «Пути эволюции животного мира» и книгу Д. М. Федотова (1966) «Эволюция и филогения беспозвоночных животных».
Несколько слов здесь уместно сказать о некоторых соображениях и принципах, которыми мы руководствовались в нашей работе.
При установлении общности происхождения органов и других структур мы пользовались критериями гомологии, предложенными Ремане, который критически разобрал это основное в сравнительной морфологии понятие (Remane, 1956). Однако мы не можем признать удачной недавнюю попытку Гансона (Hanson, 1963) привести критерии для установления гомологии функций и процессов. Поскольку, писал Гансон, гомологичными считаются структуры, обладающие общим происхождением, мы вправе применить понятие гомологии и к физиологическим процессам, унаследованным от общих предков. Сама по себе эта мысль совершенно верна, однако на деле Гансон стремится показать, что любые две функции у разных организмов, если они достаточно сходны, являются гомологичными. Одно сходство этих функций якобы свидетельствует и о гомологии выполняющих их структур. Эти утверждения противоречат самой сущности понятия гомологии, Хорошо известно, что гомологичные органы нередко несут разные функции и что, напротив, одинаковые отправления часто свойственны не гомологичным, но лишь сходным по строению и расположению органам. Ни сходство функций, ни совпадение локализации выполняющих их структур сами по себе не имеют решающего значения для установления гомологии. Было бы излишне напоминать об этих истинах, если бы Гансон не утверждал, что одинаковое положение в теле и более или менее сходная функция сравниваемых структур вполне достаточны для доказательства их гомологии.
За существенный критерий примитивности той или иной группы мы принимаем наличие слабо специализированной и пластичной простой организации. Особенно должно привлекать в этом смысле внимание большое морфологическое многообразие, если оно наблюдается в пределах одной и той же группы сравнительно невысокого систематического ранга и затрагивает общий план строения. Наличие такой неустойчивости организации, именно большие различия в строении, числе и положении органов, характерно для очень примитивных, еще слабо специализированных групп, находящихся близ основания крупных филогенетических ветвей (Hand, 1959; Мамкаев, 1968).
По мере эволюции, происходившей в различных направлениях, как бы выкристаллизовываются все более и более устойчивые морфологические типы с более или менее ограниченными эволюционными возможностями. Если при этом все-таки имеет место значительное морфологическое разнообразие, то оно носит иной характер, так как не затрагивает уже выработанного типа организации, проявляясь в усложнении и специализации вполне сложившихся аппаратов или осуществляясь за счет развития новых органов (см.: Мамкаев, 1968). Эти представления восходят к взглядам Копа (Соре, 1887) и Депере (1915) о возникновении прогрессивных форм от неспециализированных предков и о специализации филогенетических ветвей. Сходное понимание эволюционного развития получило широкое признание(см., например: Шмальгаузен, 1938, 1939, 1969); на него оцирается,
в частности, учение об олигомеризации гомологичных органов (Догель, 1954).
По мнению В. А. Догеля, новые органы развиваются у примитивных форм, появляясь в большом числе; они слабо развиты, весьма однородны и часто располагаются беспорядочно (принцип множественной закладки новообразующихся органов). По мере дифференциации они приобретают определенную локализацию, а количество их при этом уменьшается, становясь, наконец, более или менее постоянным для данной группы животных. Вслед за В. А. Догелем мы считаем, что явление олигомеризации имеет существенное значение в решении многих вопросов филогении. По множественному или уже олигомеризованному состоянию органов нередко удается судить как о степени их примитивности, так и о сравнительной давности их возникновения, а по комбинации органов разного филогенетического возраста нередко можно сделать заключение о происхождении данной группы. Явление олигомеризации также позволяет говорить о направлении филогенетического развития. Ниже мы используем проявление принципа множественной закладки гомологичных органов и их олигомеризации главным образом при анализе эволюции органов кожного вооружения и нервного аппарата турбеллярий.
Изучая современных представителей какой-либо группы, зоолог-филогенетик стремится обнаружить среди них наиболее примитивные формы, которые можно было бы рассматривать как наиболее близкие к общим родоначальникам всей группы. Однако примитивных во всех отношениях животных не существует. В организации почти любого животного наряду с действительно примитивными чертами всегда имеются более или менее глубокие вторичные изменения. Истина, что в природе, нет живых схем и что они никогда не существовали, к сожалению, часто забывается. Неравномерность темпов эволюции разных органов — факт давно известный и привлекавший к себе пристальное внимание ряда исследователей (Шмальгаузен, 1938, 1939; Ливанов, 1955; Воронцов, 1963; Лукин, 1964). В. А. Догель, отмечая это явление, писал: «Важным моментом, определяющим строение животного, является известный консерватизм. . . образований, удерживающихся нередко еще в течение долгого времени после того, как изменение окружающих внешних условий или модификация общего плана строения требует их перестройки в определенном направлении. Именно это обстоятельство приводит к тому, что разные системы органов сплошь и рядом находятся на разных ступенях сложности строения у одного и того же животного. Идеальный случай требует для типичного целомического животного открытых в полость тела метанефридиев. Между тем у многих Polychaeta еще удерживаются протонефридии» (1940, стр. 466).
Не представляют исключения и турбеллярии. У них мы нередко обнаруживаем отдельные очень примитивные признаки, характерные для низших групп, но также спорадически рассеянные в относительно высокоразвитых группах. Таковы эпители- ально-мускульные элементы покровов, статоцист, интраэпите- лиальная нервная система, синцитиальное состояние кишечного эпителия, общий гермафродитный зачаток гонад и некоторые другие. Было бы явной методической ошибкой признать эти признаки вторичными упрощениями на том основании, что они встречаются в виде изолированных примеров в высших группах. Правда, в ряде случаев «запоздалое» проявление примитивной особенности в действительности может оказаться вторичным упрощением, вызванным, например, редукцией путем отпадения конечных стадий органогенеза или фетализации или даже спонтанного атавизма. Однако это не дает нам права приписывать такую же вторичную природу сходным признакам у явно примитивных групп. Разумеется, каждый конкретный спорный случай
такого рода должен анализироваться отдельно, с учетом наиболее вероятных филогенетических связей между сравниваемыми группами. В большинстве случаев при непредвзятом подходе этого достаточно, чтобы убедиться, что «запоздалый» примитивный признак в высшей группе кажется изолированным лишь на первый взгляд, но в действительности унаследован от далекого общего предка.
В работе филогенетика большое значение имеет реконструкция обобщенной схемы организации, характерной для данной группы животных. Для получения такого общего плана строения, «морфологического типа» или «прототипа» группы используются наиболее характерные и примитивные черты организации. Понятие о морфологическом типе относится к старинным представлениям в морфологии и зоологии; его методологическое значение и история до Дарвина и в последарвиновское время прекрасно разобраны в интересных книгах И. И. Канаева (1963, 1966). Здесь уместно отметить лишь следующее. Хотя морфологический тип не воплощается реально ни в какой конкретной живой форме, так как в нем учитываются лишь общие примитивные черты и отброшены все индивидуальные и вторичные признаки, сам он тем не менее представляет собой вполне объективную реальность, поскольку все существенные его элементы в их характерных сочетаниях действительно имеются у разных представителей данной группы.
Классическим примером последовательного применения в сравнительной анатомии и филогенетике метода морфологического типа может служить воссоздание генеалогии низших позвоночных академиком А. Н. Северцовым (1916, 1934, 1967). Им реконструирована серия морфологических типов, дающих очень полное представление о главнейших этапах филогенетического развития позвоночных, — это общие планы строения первичных Acrania, гипотетических Protocraniata, Entobranchia и Ectobranchia, Proto- cyclostomata. и Protognathostomata. В области сравнительной анатомии беспозвоночных также нет недостатка в подобных примерах реконструкции морфологического типа отдельных групп, которые обычно рассматриваются и как обобщенные схемы пред- ковых форм. Достаточно, например, вспомнить геккелевскую Gastraea, или мечниковскую Phagocytella, или гипотетического предка моллюсков Prorhipidoglossum, представление о котором было блестяще разработано Пельзенером (Pelseneer, 1906).
Одну из попыток воссоздания морфологического типа класса Turbellaria уже сделали Рейзингер (Reisinger, 1928—1933) и Акс (Ах, 1961), причем последний исходил из представлений о регрессивной эволюции плоских червей, связанных с теорией Spiralia. Мы не разделяем этих представлений и ставим перед собой задачу реконструировать морфологический тип турбеллярий, основываясь на положениях ацельной теории происхождения ресничных червей, разработанной Граффом.
Среди многочисленных точек зрения на природу турбеллярий многие широко известны и уже обсуждались достаточно подробно в советской зоологической литературе. Таковы ктенофорная гипотеза Ланга, подвергнутая критике В. Н. Беклемишевым (1944, 1964), и гипотеза целлюляризации Хаджи, Штейнбека и Гансона, детально рассмотренная недавно А. В.Ивановым (1968). Другие концепции не разбирались еще в отечественной литературе. Мы имеем в виду прежде всего взгляды Ремане и его сторонников, а также высказывания В. А. Догеля о полифилетическом происхождении турбеллярий и филогенетические соображения В. Н. Беклемишева и Карлинга. Им мы уделим особое внимание.
СОСТАВ И ХАРАКТЕРИСТИКА PLATHELMINTHE S
Вслед за большинством современных авторов мы рассматриваем плоских червей как самостоятельный тип низших Proto- stomia. В состав Plathelminthes, по нашему мнению, должны входить следующие классы:
Xenoturbellida 4. Monogenoidea
Turbellaria 5. Gyrocotyloidea
Trematoda 6. Cestoidea.1
Так называемых Mesozoa (Orthonectida и Dicyemida) мы в противоположность В. А. Догелю (1959) и В. Н. Беклемишеву (1964) отказываемся причислять к плоским червям. Мы присоединяемся к Бресслау и Рейзингеру (Bresslau u. Reisinger, 1928— 1933), отнесшим Temnocephalida к Turbellaria в качестве отдельного отряда на основании большой близости их к Neorhabdocoela. Далее мы полагаем, что и Udonellida представляют собой отряд Turbellaria Neoophora. Этих немногочисленных и своеобразных комменсальных плоских червей с железистой присоской на заднем конце тела (рис. 1), обитающих на паразитических копепо- дах Caligidae, издавна считают моногенеями (см., например: Sproston, 1946; Hyman, 1951; Baer et Euzet, 1961). В 1952 г.
А. В. Иванов, детально изучив организацию Udonella caligorum, выделил их в отдельный класс плоских червей, который, по его мнению, произошел вполне самостоятельно от Neorhabdocoela. Основаниями для этого послужили главным образом отсутствие у удонеллид личинки, снабженной церкомером с его хитиноид- ным вооружением, и очень своеобразное строение протонефри- диального аппарата. Класс Udonelloidea был признан многими зоологами (Догель, 1954, 1959; Быховский, 1957; Беклемишев, 1964; Гинецинская, 1968; Шульц и Гвоздев, 1970). Однако теперь мы думаем, что отсутствие церкомера, не позволяющее относить
удонеллид ни к Monogenoidea, ни вообще к Cercomeromorphae (см.: Быховский, 1937, 1957), не говорит, разумеется, против близкого их родства с Neorhabdocoela, с которыми они сходны по плану строения. Вместе с тем особенности строения выделительного аппарата удонеллид (см. стр. 93), несмотря на все его
своеобразие, вряд ли можно считать до- статочными для обособления их в само- г стоятельный класс.
/ I Gnathostomulida (рис. 2, А) мы также
f
?:■
■■■: \
г рассматриваем только как отряд
Turbellaria
Archoophora.
Их
план строения прекрасно укладывается
в рамки морфологического типа
турбеллярий: кожно-мускульный мешок,
нервная система, органы чувств, кишечник,
тип глотки и половая система ничем
принципиальным не отличаются от
таковых турбеллярий. Для гнатостомулид
характерно примитивное квартетное
спиральное дробление. Немногие
оригинальные черты организации
гнатостомулид заключаются в развитии
жгутикового, а не ресничного эпидермиса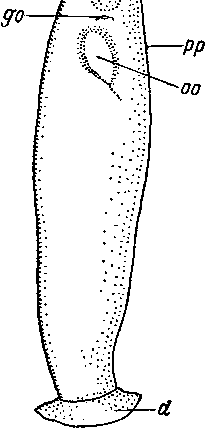
Рис. 1. Udonella caligorum (Udonellida). Внешний вид с брюшной стороны. (По: Иванов, 19526).
d — прикрепительный железистый диск, g — железистая ямка, go — половое отверстие, о — ротовое отверстие, оо — просвечивающее яйцо, ph — просвечивающая глотка, рр — выделительное отверстие.
(рис. 8), хитиноидных подвижных челюстей в глотке (рис. 2, Б) и, наконец, в своеобразной форме спермиев у некоторых представителей (рис. 3, Б). Однако первая и последняя особенности не являются даже органологическими признаками и не имеют поэтому серьезного таксономического значения, хотя, разумеется, жгутиковый эпидермис сам по себе чрезвычайно архаичный признак. Что касается челюстей, то развитие подобного рода хватательных хитиноидных образований, хотя и не связанных с глоткой, свойственно некоторым турбелляриям (хитиноидные «челюсти» и крючки в хоботке некоторых Neorhabdocoela Kalyp- torhynchia). Все перечисленные оригинальные черты Gnathostomulida недостаточны для выделения их в самостоятельный класс и тем более тип, как это делают некоторые авторы (Ах, 1960, 1966; Riedl, 1969).
Единственный представитель группы Xenoturbellida — Хепо- turbella bocki — был открыт Вестбладом довольно давно (Westblad, 1949а). Несмотря на это, систематическое положение Xenoturbellida остается неясным.Xenoturbella имеет очень простое строение, й простота эта скорее всего первична (рис. 4). Эту форму можно рассматривать как очень примитивную турбеллярию, что и делал описавший ее Вестблад. Этой же точки зрения придерживались В. Н. Беклемишев (1952, 1963а, 1964) и А. В. Иванов (1968). Действительно, ряд особенностей в организации Xenoturbella сходен с таковым турбеллярий, а именно общий ресничный покров, кожномускульный мешок, паренхима, вентральный рот с зачаточной глоткой, мешковидный кишечник, кожный нервный плексус, статоцист на переднем конце тела, гермафродитизм и гонады ацелоид- ного типа. Архитектонических различий между Xenoturbellida и Turbellaria нет.
Однако
Акс подверг это мнение серьезной критике,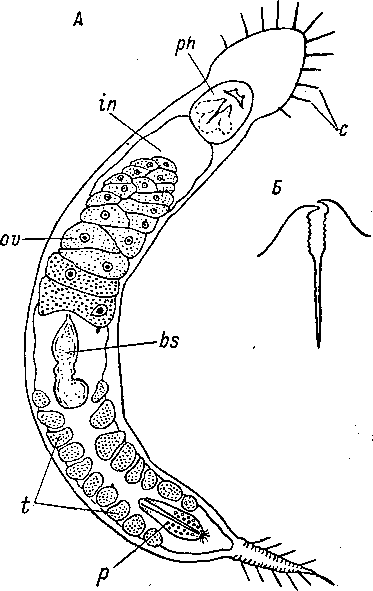
Рис. 2. Gnathostomula paradoxa (Gnathostomulida).
А — внешний вид и внутреннее строение, Б — челюсти. (По: Ах,
1956). Ьз — семенная сумка, с — осязательные щетинки, in — кишка, ph — глотка с базальной пластинкой и челюстями, ov — яичник, р — мужской совокупительный орган, t — семенник.
заявив не без основания, что, во-первых, все признаки Xenoturbella, общие с турбелляриями, широко распространены среди низших беспозвоночных и что, во-вторых, Xenoturbella свойственны некоторые «специфические качества», отсутствующие у турбеллярий (Ах, 1960, 1961, 1963а). Эти особые качества были обнаружены Рейзингером при дополнительном изучении животного (Reisin- ger, 1960). Оказалось, что эпидермис Xenoturbella (рис. 5, А) отличается от кожного эпителия турбеллярий своей многослой- ностью, составом из разнородных клеток и чрезвычайно напоминает таковой Enteropneusta (рис. 5, Б). Кроме того, благодаря своему эпидермальному положению статоцист Xenoturbella скорее сходен с аборальным органом гребневиков, чем со статоци- стом турбеллярий. Но более всего, по Рейзингеру, орган равновесия напоминает статоцисты некоторых голотурий (Synaptidae
и klpidiidae), стенки которых тоже состоят из многих клеток и которые содержат, как и у Xenoturbella, несколько (до 20) литоцитов (рис. 6, А). Наконец, спермин Xenoturbella (рис. 6, Б) принадлежат к обычному примитивному типу сперматозоидов,
Рис.
3. Спермин Gnathostomulida.
А
— Pterognathia
sp.
с
живого экземпляра, В
— Gnatkostomula
axi
с
электронномикроскопической фотографии
(А — по: Sterrer,
1969;
Б
— по: Graebner,
1969).
широко распространенному у Metazoa, в том числе у Deuterosto- mia, тогда как у турбеллярий нитевидные спермии вторично изменены. Поэтому Рейзингер и Акс решительно возражают против включения Xenoturbellida в класс Turbellaria и высказываются за их родство с Deuterostomia.
Основания для такого вывода очень шатки. Все перечисленные признаки, указывающие якобы на близость к Deuterostomia,
носят чисто гистологический характер и не могут иметь большого таксономического значения. Эпидермис Xenoturbella относится к типу немертинового эпидермиса, который, кроме Enteropneusta, развит также у Nemertoderma (рис. 11), у Aechmalotus (Turbellaria, Acoela) и у некоторых немертин. Этот тип эпидермиса возникает явно независимо в разных группах. Сходство статоцистов Xenoturbella и голотурий несомненно носит случайный характер и есть результат конвергенции, поскольку статоцисты Xenoturbella и турбеллярий по меньшей мере обладают общей гомологией (гомоплазией), ибо в обоих случаях это органы равновесия на переднем анимальном полюсе, имеющем тенденцию у низших Metazoa к развитию чувствительных приспособлений (см.: Беклемишев, 1964), тогда как у голотурий они никакого отношения к этим первичным ани- мальным органам чувств не имеют. Хотя между статоцистами Xenoturbella и голотурий имеется гистологическое сходство, гораздо важнее, что ни по числу, ни по положению
Рис.
4. Xenoturbella
bocki
(Xenoturbellida).
А
— внешний вид с брюшной стороны, В
— сагиттальный разрез. (По: Westblad,
1949а).
ер
— эпидермис, in
—
кишка, п
—нервный слой, о — ротовое отверстие,
оо — ооциты, г — поперечная ресничная
бороздка, si
—
статоцист.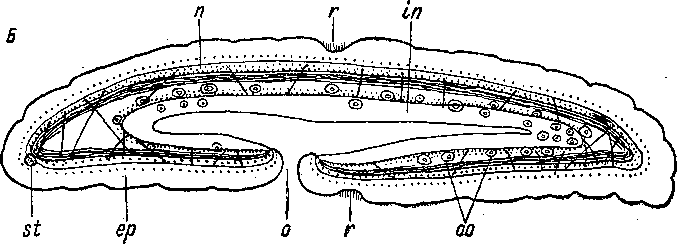
между
ними нет ничего общего. Статоцисты
Synaptidae
расположены
по два по бокам от каждого радиального
нервного ствола, в том месте, где он
покидает окологлоточное известковое
кольцо, т. е. их 10, и они имеют радиальное
расположение — каждая пара есть элемент
одного из пяти радиусов. У Elpidiidae
число
органов равновесия значительно больше,
а расположение их своеобразнее. Оба
радиальных нерва бивиума, т. е. спинной
стороны, несут по два статолита. Средний
нерв тривиума совсем лишен этих органов,
зато боковые его нервы имеют каждый от
26 до 40 статоцистов. Последние у голотурий
отличаются и тем, что лежат 
в соединительнотканном слое кожи, т. е. полностью отделены уже от эпидермиса (см.: Догель, 1940). «Независимое возникновение статоцистов в разных группах животных, — пишет В. Н. Беклемишев (1964, II, стр. 139), — вытекает из различного их размещения». Следовательно, нет оснований искать гомологию
Рис.
5. Эпидермис Xenoturbellida
и
Enteropneusta.
(Из:
Reisinger,
1960).
А
— Xenoturbella
bocki,
В
— Balanoglossus
sp.
bm
—
базальная мембрана, ер
—• эпидермальные клетки, g
—
железистые клетки различного типа, gc
—
нервная клетка, п
— нервный слой, sc
—
чувствительная клетка.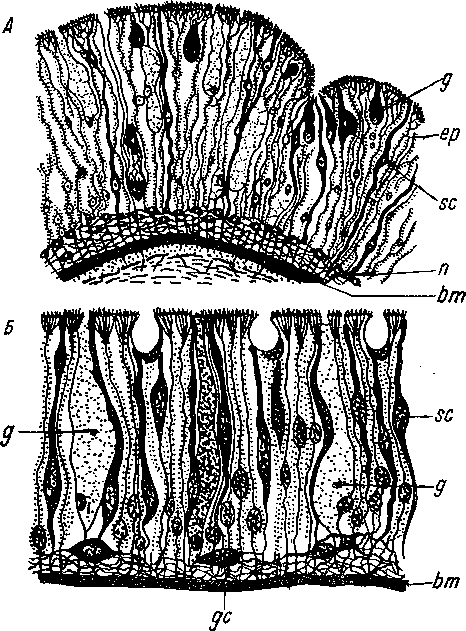
между статоцистами Xenoturbella и голотурий, но ничто не говорит против гомологизации статоцистов Xenoturbella и Turbellaria.
Отличия спермиев Xenoturbella от таковых турбеллярий столь же существенны (хотя на этот раз речь идет о цитологических различиях). У турбеллярий спермии далеко не однотипны. Так, очень резко отличаются сперматозоиды Gnathostomulida, которые у одних родов обнаруживают типичное для «нормального»
спермин строение (Pterognathia — рис. 3, А), у других же (Gna- thostomula — рис. 3, В), как показывает электронный микроскоп, имеют вид округлых клеток без жгутов, но с пучками многочисленных «ножек» (Graebner, 1969; Sterrer, 1969). Во всяком случае примитивный тип спермиев у Xenoturbellida указывает на существование у них наружного оплодотворения; в пользу этого предположения говорит и отсутствие мужского совокупительного органа.
В
Рис.
6. Xenoturbella
bocki
(Xenoturbellida).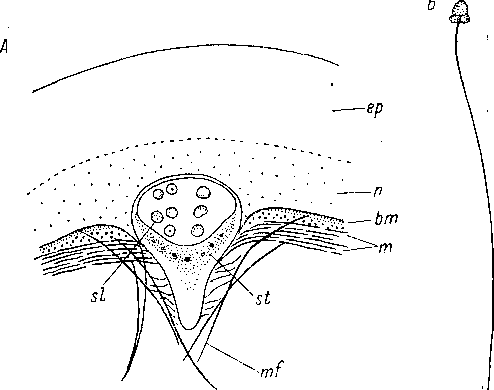
А — горизонтальный разрез через статоцист, В — спермий. (По: Westblad, 1949а). Ьт — базальная мембрана, ер — внешняя часть эпидермиса, т. — мускульные слои, mf — мускульные волокна, п — нервный слой, si — статоцист,
si — литоциты.
Таким образом, Xenoturbellida очень близки к низшим тур- белляриям, и в сущности довольно безразлично, вводить ли их в состав Turbellaria или ставить рядом с ними в качестве самостоятельного класса. Однако в первом случае пришлось бы расширять диагноз класса Turbellaria до каких-то неопределенных границ.
Иегерстен (Jagersten, 1959) рассматривает Xenoturbellida как реликтовую группу, стоящую ближе всего к гипотетической Bilaterogastraea, которую он считает общим предком всех Metazoa. Ремане (Remane, 1959), Д. М. Федотов (1966) и Ю. В. Мамкаев (1967) отказались помещать Xenoturbella среди турбеллярий.
К сожалению, Xenoturbella bocki еще недостаточно изучена, ее эмбриональное развитие и жизненный цикл неизвестны. Было бы преждевременным делать окончательные выводы. Мы
А. В. Иванов и Ю. В. Мамкаев
17
склонны нровизорно рассматривать Xenoturbella как представителя самостоятельного класса в составе Plathelminthes. Что касается предположения Рейзингера о неотенической природе Xenoturbella, то оно остается чисто умозрительным, в защиту его нельзя привести никаких существенных доводов.
Диагноз Plathelminthes, по нашему мнению, должен иметь следующий вид.
Низшие несегментированные протаксонные Protostomia без полости тела. Дефинитивный рот гомологичен бластопору. Тело одето ресничным (в редких случаях жгутиковым) эпидермисом, однако у многих форм ресничный покров редуцирован. Кишечник у примитивных представителей отсутствует, у большинства слепо замкнут, или полностью редуцирован. Между стенкой тела и внутренними органами развита паренхима. Имеется кожномускульный мешок. Выделительные органы отсутствуют или представлены протонефридиями. Нервный аппарат—-от кожного плексуса до высокоразвитого мозга с ортогональными стволами. Кровеносная и дыхательная системы отсутствуют. Гермафродиты большей частью с внутренним оплодотворением. Дробление спиральное, часто вторично измененное.
ОБЩАЯ ХАРАКТЕРИСТИКА TURBELLARIA
Turbellaria — наиболее примитивный, основной класс Plathelminthes, исходный для всех остальных плоских червей. Общая его характеристика связана со значительными трудностями, во-первых, из-за необычайно большого морфологического разнообразия турбеллярий и, во-вторых, потому, что близость ресничных червей к соседним классам иногда столь велика, что подчас бывает нелегко провести границу. Относительно некоторых групп трудно решить, следует ли ввести их в состав Turbellaria или придать им ранг класса. Таковы прежде всего Xenoturbellida, которых многие причисляют к турбелляриям. Тетпосе- phalida и Udonellida нередко выделяются в качестве самостоятельных классов. Даже некоторые Nemathelminthes, именно Gastro- tricha, еще чрезвычайно близки к Turbellaria. В результате, пожалуй, нельзя указать ни одного признака, который был бы обязательным для всех членов класса Turbellaria и в то же время отсутствовал у всех остальных Plathelminthes.
При сравнении Turbellaria с соседними классами нетрудно заметить разительный контраст между их огромным морфологическим разнообразием и относительной стабильностью в организации остальных плоских червей. Несомненно, что само разнообразие планов строения турбеллярий следует считать одной из самых характерных особенностей этого класса, свидетельствующей о его общей примитивности.В характеристике Turbellaria, как нам представляется, должны быть отмечены следующие их черты.
Это в основном свободноживущие хищные Plathelminthes, лишенные церкомера, с простым жизненным циклом. Тело одето мерцательным покровом. Положение рта варьирует в широких пределах — от заднего конца тела до переднего полюса. Обычно имеется глотка — простая, складчатая, массивная или конусовидная. Кишечник у примитивных форм не развит, и пищеварение совершается в центральной паренхиме, или имеется более или менее стабильная эпителизованная кишка. Органы чувств представлены осязательными сенсиллами, обонятельными ямками, статоцистом и глазами. Половая система чрезвычайно разнообразного строения. У примитивных форм женские гонады представлены только яичниками, производящими энтолецитальные яйца, которые испытывают спиральное дробление; у высших форм женские гонады в виде гермовителляриев или гермариев и жел- точников, яйца экзолецитальные, с измененным типом дробления. Всегда имеется мужской совокупительный орган; женские совокупительные органы у наиболее примитивных форм не развиты.
Турбеллярии обитают преимущественно в морях, но имеется много пресноводных и немало наземных форм. Некоторые турбеллярии ведут паразитический или комменсальный образ жизни. Известно свыше 3000 видов.
Размеры турбеллярий невелики. Большинство видов имеет от 0.4 до 5 мм в длину, но поликладиды и наземные планарии отличаются значительной величиной. Самая большая турбелля- рия Bipalium javanum достигает 60 см в длину. Многие поликладиды и наземные планарии^отличаются яркой^окраской.
КЛАССИФИКАЦИЯ TURBELLARIA
Старая система Граффа, делившая турбеллярий на 5 отрядов (Acoela, Alloeocoela, Rhabdocoela, Tricladida и Polycladida), основывалась главным образом на форме кишечника. Влияние этой системы продолжалось очень долго, и еще недавно ее придерживалась Гайман (Hyman, 1951). Серьезный шаг вперед сделал Мейкснер (Meixner, 1926), который, во-первых, выделил из гетерогенного комплекса Rhabdocoela низшие формы, придав им ранг отрядов (Catenulida и Macrostomida), и, во-вторых, присоединил Tricladida к отряду Alloeocoela. В самом деле, лишенные желточников катенулиды и макростомиды отличаются примитивностью строения, тогда как обладающие желточниками рабдо- целиды представляют собой наиболее дифференцированные формы. Последних Мейкснер обозначил как отряд Neorhabdocoela. Что касается Tricladida, то они мало чем отличаются от Proseriata, входящих в состав Alloeocoela. Таким образом, Мейкснер различал 6 отрядов: Acoela, Catenulida, Macrostomida, Polycladida, Alloeocoela и Neorhabdocoela.
Дальнейшие улучшения в систему турбеллярий внес Карлинг (Karling, 1940), который расчленил довольно искусственную группу Alloeocoela на отряды (Prolecithophora, Lecithoepitheliata и Seriata), существенно различные по строению половой системы и глотки. Он же предложил объединить Acoela, Catenulida, Macrostomida, Nemertodermatidae, Hofsteniidae и Proplicastomata 1 в низший отряд Archoophora, охватывающий турбеллярий с простыми пищеварительными и половыми органами, не имеющих жел- точников и откладывающих энтолецитальные яйца. Однако Карлинг сделал исключение для поликладид, оставив их за пределами Archoophora1, он предполагал, что отсутствие у них желточников вторично и есть результат редукции этих органов. Это утверждение представлялось недостаточно обоснованным, и вскоре Вест- блад (Westblad, 1948) решительно разделил всех турбеллярий на два отряда — Archoophora и Neoophora. Первые характеризуются отсутствием желточных клеток и энтолецитальными яйцами; к ним принадлежат все Archoophora Карлинга и Polycladida. Вторые обладают желточными клетками и экзолецитальными яйцами; к ним относятся все остальные турбеллярии.
Это расчленение класса турбеллярий на две группы, представляющее собой признание двух основных уровней организации, было воспринято разными зоологами различно.
Так, Гайман (Hyman, 1959, стр. 734) решительно отказывается рассматривать Archoophora и Neoophora как естественные группы и называет это деление турбеллярий «совершенно негодным» («unqualifiedly bad»). Другие авторы (Ах, 1956, 1961, 1963а; Riedl, 1959а; Luther, 1960; Boaden, 1963; Беклемишев, 19636) относят обе упомянутые группы не к систематическим единицам, а лишь к искусственным объединениям форм одинакового организационного уровня («Stadiengruppen»). Карлинг (Karling, 1967а), который недавно вернулся к этому вопросу, считает Archoophora естественной группой, a Neoophora — полифилети- ческим комплексом неродственных форм. Наконец, Ю. В. Мамкаев (1967) придает Archoophora и Neoophora значение подклассов.
Находя позицию Гайман необоснованной и чрезмерно крайней, мы присоединяемся к концепции двух уровней организации. По Ремане (Remane, 1956, стр. 127—128), «Stadiengruppen» объединяют в себе гетерогенные формы и распадаются, как только родственные отношения между ними становятся известными. Акс и В. Н. Беклемишев совершенно правы, подчеркивая, что обе группы объединены не по комплексу морфологических особенностей, а лишь по единичному признаку — наличию или
отсутствию энтолецитальных яиц и спирального дробления.
Вслед за ними мы полагаем, что правильнее всего в классе Turbellaria различать две ступени организации — низшую и высшую. Система Turbellaria, следовательно, принимает следующий вид.
Класс Turbellaria Ehrenberg, 1831
Отр. Acoela Uljanin, 1870 Отр. Gnathostomalida Ах, 1956 Отр. Macrostomida Meixner, 1926 Отр. Catenalida Meixner, 1926 Отр. Polycladida Lang, 1884
I. Archoophora .
Отр. Lecithoepitheliata Reisinger, 1926 Отр. Prolecithophora Karling, 1940 Отр. Seriata Westblad, 1935 (Proseriata Meixner, 1938 -)- Tricladida Graff, 1912—
1917)
II. Neoophora
Отр. Neorhabdocoela Meixner, 1938 Отр. Temnocephalida Bresslau et Reisinger, 1928—1933 Отр. Udonellida ordo nov.
ПЕРВИЧНАЯ ГИСТОЛОГИЧЕСКАЯ СТРУКТУРА TURBELLARIA
При достаточно детальном анализе строения Metazoa никаких сомнений в отношении их клеточной природы обычно не возникает, тело их всегда может быть разложено на ткани, состоящие из клеток. Это вполне естественно и согласуется с основными положениями клеточной теории, обязательными для всех Eukaryota. Однако в отношении одного из отрядов Turbellaria, именно Acoela, довольно широкое распространение получило мнение, что они, по крайней мере частично, лишены настоящего клеточного строения. Это утверждение возникло скорее как теоретический постулат, вытекающий из существа гипотезы целлюляризации (Hadzi, 1944, 1963; Hanson, 1958; Steinbock, 1958), пытавшейся выводить Acoela непосредственно из сомателл многоядерных инфузорий. Поэтому в литературе последних десятилетий большое внимание уделялось вопросу о первичной гистологической структуре бес- кишечных турбеллярий (Логачев, 1957; Hanson, 1958; Steinbock, 1958; Reisinger, 1959; Ах, 1961; Pedersen, 1964; Иванов, 1968, 1971а).
По утверждению сторонников цилиатно-ацельной теории, современные Acoela обладают плазмодиальной, неклеточной структурой. Гансон и Штейнбек допускают даже, что вторичная цел- люляризация почти не успела нарушить это первичное состояние, тогда как Хаджи считает, что многие органы и ткани (эпидермис, глотка, гаметы) уже полностью целлюляризовались.
Вслед за Вестбладом (Westblad, 1948) многие авторы стали различать у Acoela «эпицитий» — поверхностный эпидермальный симпласт, «эктоцитий» — лежащий глубже синцитиальный слой паренхимы и «эндоцитий» — центральную часть паренхимы. В отрицании клеточного строения у Acoela особенно далеко зашел Штейнбек (Steinbock, 1952, 1958), утверждавший, что так называемый первичный плазмодий Acoela есть не только примордиальная ткань турбеллярий, но и первичная ткань — «Archi- histon вообще всех Metazoa». По Штейнбеку, из первичного плазмодия образовались клетки Metazoa, а из последних — клеточные слои тела — экто-, мезо-.и энтодерма, которые, следовательно, гомологичны плазмодиальным слоям эпи-, экто- и эндо- цитию.
Однако учение о первичности плазмодиальной структуры находится в противоречии с клеточной теорией и опровергается твердо установленными гистологическими, эмбриологическими фактами и экспериментами, а в последнее время и электронномикроскопическими работами.
Интересные аргументы против гипотезы первичного плазмо- диального строения Metazoa привел Рейзингер (Reisinger, 1959). По его наблюдениям, у пресноводной турбеллярии Otomesostoma кишечник становится синцитиальным при проникновении в его стенку кокцидий, но после изгнания паразитов вновь приобретает клеточную структуру. У хоботковых турбеллярий Kalypto- rhynchia Рейзингер добился распадения нормально синцитиального эпидермиса на клеточные территории, применив чисто механическое раздражение. Можно поэтому согласиться с Рей- эингером (Reisinger, 1961, стр. 9), утверждающим, что «синцитий представляет собой только структурно-динамическое состояние скрытого, но всегда существующего клеточного строения ткани».
Что же касается Acoela, то представление об их исключительной синцитиальности просто не отвечает действительности. Давно известно, что при микроскопировании in vivo, а также при изучении окрашенных срезов, приготовленных после соответствующих фиксаций, клеточные границы обнаруживаются почти во всех тканях и органах бескишечных турбеллярий (Jensen, 1878; De- lage, 1886; Bohmig, 1895, 1905; Graff, 1904—1908; Luther, 1912; Беклемишев, 1915; Иванов, 1952a). Нет смысла перечислять здесь все твердо установленные примеры клеточного строения у Acoela. Недавно подробные обзоры их были сделаны Аксом (Ах, 1961, 1963а) и А. В. Ивановым (1968). Сейчас можно уже не сомневаться, что бблыпая часть органов Acoela имеет нормальное клеточное строение. Большинство современных зоологов признает только вторичное исчезновение клеточных границ в некоторых тканях Acoela (Беклемишев, 1937, 1964; Ливанов, 1945; Иванов, 1952а, 1968; Мамкаев, 1967). По Беклемишеву, у бескишечных турбеллярий встречаются почти все типы клеточного эпидермиса,
свойственные беспозвоночным, но все они могут существовать в двух вариантах — клеточном и синцитиальном, причем последний всегда есть производное первого.
Когда речь идет о синцитиальности паренхимы Acoela, то имеется в виду ее протоплазматическая многоядерная масса. Световой микроскоп не позволяет открыть в ней границы клеточных территорий. Некоторые ее свойства недавно были изучены экспери-" ментально у Convoluta convoluta (Мамкаев и Серавин, 1963). Она отличается полужидкой консистенцией, изолированные ее участки сохраняют амебоидную подвижность в морской воде в течение нескольких часов.
Электронномикроскопические исследования обнаружили клеточную природу паренхимы Acoela. По' обстоятельным наблюдениям Педерсена (Pedersen, 1964), тело Convoluta convoluta слагается исключительно из клеток. Хорошо прослеживаются контуры эпидермальных и мышечных клеток и клеток периферической паренхимы. Многочисленные цитоплазматические мембраны обнаруживаются и в центральной паренхиме. Эти данные подтверждаются и другими работами (см.: Dorey, 1965).
В клеточной организации Acoela мы и сами могли убедиться в начатых нами электронномикроскопических исследованиях Convoluta convoluta и Anaperus biaculeatus. Все эти факты свидетельствуют против плазмодиальной природы бескишечных турбеллярий и выбивают почву из-под ног защитников гипотезы целлюляризации.
По мнению Педерсена, центральная паренхима Convoluta convoluta слагается из очень крупных амебоидных, сильно ветвящихся клеток, многочисленные отростки которых переплетаются друг с другом. Они ограничены клеточной мембраной и, помимо ядра, содержат митохондрии, аппарат Гольджи, рибосомы, эндо- плазматическую сеть канальцев и гранулы запасных питательных веществ. Кроме того, в клетках имеются многочисленные пищеварительные вакуоли. Несколько иначе Педерсен описывает гистологическое строение небольшой части центральной паренхимы, способной выступать наружу через ротовое отверстие. Здесь обычная клеточная структура легко нарушается, причем может возникать временное обратимое синцитиальное состояние.
Ю. В. Мамкаев (1967) допускает, что и в остальной части пищеварительной паренхимы клеточные мембраны находятся в очень лабильном состоянии и могут легко формироваться и столь же легко исчезать, причем возникают быстро преходящие синцитиальные структуры. Поэтому центральную паренхиму можно трактовать как подвижный конгломерат клеток, представляющий *■
собой очень неустойчивую форму клеточной организации. Эти заключения Мамкаев подкрепляет ссылкой на тот хорошо известный факт, что клеточные пограничные мембранные структуры вообще способны легко трансформироваться и, в частности, могут
давать внутренние цитоплазматические мембраны (см., например: Wohlfarth-Bottermann, 1961, 1964; Робертсон, 1964).
В дальнейшем во всех случаях, когда нам придется пользоваться термином «синцитий», в том числе применительно к центральной пищеварительной паренхиме, мы всегда будем иметь в виду его вторичное происхождение.
Подводя итоги, мы можем сделать вывод, что вопреки утверждениям сторонников теории целлюляризации бескишечные турбеллярии по своей клеточной природе ничем не отличаются от всех остальных Metazoa. Структура их тканей не дает никаких свидетельств в пользу гипотезы происхождения турбеллярий от инфузорий. Отсутствие клеточных границ в эпидермисе некоторых видов Acoela (которое, может быть, подтвердится в дальнейшем) носит явно вторичный характер и указывает на наличие синцития, а не первичного плазмодия. Из этого, однако, вовсе не следует, что организация Acoela и в других отношениях подверглась вторичным изменениям, что ее бросающаяся в глаза простота есть результат вторичных упрощений, как полагают многие выдающиеся зоологи (Ливанов, 1945; Remane, 1954; Marcus, 1958; Brien, 1961, 1969, и др.). С другой стороны, было бы неправильно и несправедливо считать, как это делает Акс, что решительно все концепции, рассматривающие состояние тканей у бескишечных турбеллярий как примитивное, вообще не согласуются с фактом клеточного строения турбеллярий.
Мы придерживаемся той точки зрения, что именно Acoela — самая примитивная группа турбеллярий. Это представление берет свое начало от И. И. Мечникова (1877а, 18776, 1886) и от Граффа (Graff, 1882, 1891, 1904—1908, 1904) и защищалось многими зоологами, лично изучавшими Acoela (Bresslau, 1928—1933; Reisinger, 1928—1933; Беклемишев, 1937, 1964; Meixner, 1938; West- blad, 1948; Hyman, 1951; Иванов, 1952a, 1971a; Мамкаев, 1967). Ниже мы постараемся доказать это, здесь же отметим, что совершенно особый теоретический интерес представляют первичные гистологические особенности у Acoela, в частности строение их паренхимы.1В этом отношении они отличаются от всех остальныхEumetazoa, в том числе и от других турбеллярий.
Наиболее глубоким анализом организации Acoela мы обязаны
В. Н. Беклемишеву (1944, 19636, 1964). При изучении строения низших Metazoa он опирался на развитую им концепцию о смешанной паренхиме как о наиболее примитивном структурном состоянии многоклеточных животных.
Смешанная паренхима еще не является истинной тканью, это — соединение разнородных клеток, несущих самые различные функции, равномерно перемешанных, не собранных еще в однородные клеточные комплексы (Беклемишев, 1925). «Смешанно паренхимная структура в чистом своем виде не встречается ни у кого из взрослых Metazoa, но существуют две группы животных, структуры которых к ней приближаются: это губки и бески- шечные турбеллярии», — пишет В. Н. Беклемишев (1964, II, стр. 5—6).
Действительно, в паренхиме Acoela перемешаны самые разнородные клетки (рис. 7): соединительнотканные клетки различного типа, камбиальные, пищеварительные разветвленные и блуждающие фагоциты; среди них попадаются железистые и половые клетки. Миоциты пронизывают паренхиму в разных направлениях и пересекают скопления половых клеток и мозг. Сквозь паренхиму \ тянутся скопления клеток подкожного нервного сгущения. Половые клетки — сперматогонии и овогонии — лежат в паренхиме среди других клеточных элементов и только у взрослых животных образуют более или менее компактные скопления.
Однако «чистота» смешанной паренхимы у Acoela нарушена развитием у них наружного клеточного слоя — эпидермиса, который несомненно уже может быть назван эпителиальной тканью. Впрочем, эпидермис иногда очень мало обособлен от паренхимы, как это имеет место в случае очень примитивного эпидермиса Oligochoerus erythrophthalmus, и тогда он составляет как бы только поверхностный слой паренхимы.
Клеточные элементы в паренхиме Acoela не образуют еще комплексов, заслуживающих наименования тканей. Тем не менее в их расположении имеется уже определенная правильность архитектонического характера. Отдельные клетки расположены обычно вполне целесообразно в отношении целого организма. Так, пищеварительные клетки преобладают в центральной области тела, где образуют пищеварительную паренхиму, опорные соединительнотканные и камбиальные элементы сосредоточены в периферическом слое паренхимы, мускульные клетки расположены не как попало, а соответственно тем функциям, которые они выполняют. Нервные клетки сосредоточены на переднем конце тела, мужские половые клетки — на спинной, а женские — на брюшной стороне.
Изложенная здесь трактовка структуры Acoela, предложенная
В. Н. Беклемишевым, представляется нам совершенно справедливой. Мы не можем поэтому не согласиться с ним в том, что архитектоническая упорядоченность клеток в теле бескишечных турбел-
Яярий есть проявление некоторой тенденции к сегрегации, т. ё. к обособлению и собиранию воедино клеток одной системы. Иначе
Рис.
7. Convoluta
sordida
(Acoela).
Часть
поперечного разреза в области рта. (По:
Graff,
1904—1908).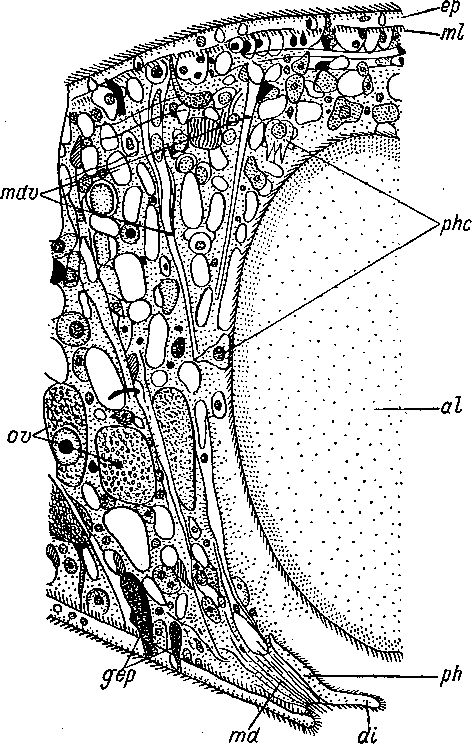
al — заглоченная добыча, ер — эпидермис, эпителиальная пластинка, содержащая часть ядер, другая часть ядер погружена в паренхиму, di — диафрагма края рта, gep — кожные железы, md — мускулы-дилататоры рта, mdv — дорсо-вентраль- ные мускульные волокна, ml — продольные мускульные волокна, ph — глотка, phc — пищеварительные клетки, фагоциты, ov — ооциты.
говоря, у Acoela мы видим зачаточное тканеобразование. У других турбеллярий этот процесс продвинулся значительно дальше: их паренхима, обычно уже резко обособленная от кишечного эпителия, представляет собой хотя и несколько разнородную по клеточному составу и мультифункциональную, но по существу уже настоящую соединительную ткань.
Вопреки мнению многих современных зарубежных зоологов, которые, будучи в большинстве своем последователями Геккеля, принимают эпителий за исходную гистологическую структуру Metazoa, мы вслед за В. Н. Беклемишевым (1925, 1964) считаем, что паренхима представляет собой наиболее примитивную ступень развития ткани. Признав геккелевскую Гастрею с ее двумя эпителиальными зародышевыми слоями за исходную форму многоклеточных, эти исследователи тем самым выпускают из поля зрения целый этап в эволюции — период паренхиматозной организации и переход от нее к организации более высокой, характеризующейся все возрастающей эпителизацией.
СРАВНИТЕЛЬНО-АНАТОМИЧЕСКИЙ ОБЗОР TURBELLARIA
Интересные и содержательные обзоры организации ресничных червей в последние десятилетия были сделаны В. Н. Беклемишевым (1944, 19636), Н. А. Ливановым (1945, 1970), Гайман (Hyman, 1951), Аксом (Ах, 1961, 1963а), Бошаном (Beauchamp, 1961) и Д. М. Федотовым (1966). Сопоставление их весьма поучительно, так как они осуществлены на основе разных теоретических представлений и приводят к различным заключениям. Сравнительноморфологический анализ турбеллярий должен дать объективные основания как для построения их естественной системы, так и для создания морфологического прототипа всего класса.
Покровы и кожно-мускульный мешок
Эпидермис
Эволюция покровов у турбеллярий полнее всего была изучена
В. Н.. Беклемишевым (1925, 19276, 1937, 1944), и мы будем опираться в основном на его выводы, которые представляются нам наиболее вероятными. Исходным типом эпидермиса у ресничных червей Беклемишев считает примитивный ресничный кожный эпителий бескишечной турбеллярии Oligochoerus erythrophthalmus, описанный ниже. Однако с тех пор как открыт отряд Gnatho- stomulida, стало ясно, что прототипом эпидермиса должен считаться жгутиковый эпителий. Гнатостомулиды — единственная группа Metazoa, представители которой во взрослом состоянии одеты сплошным покровом не ресничек, но жгутов (рис. 8), выполняющих локомоторную функцию (Ах, 1956,1963b).1Сплошной жгути-
ковый эпидермис развит лишь у личинок различных беспозвоночных: паренхимул и амфибластул губок, паренхимул и планул книдарий, у личинок некоторых форонид и у бластулообразных личинок иглокожих и ланцетника (Conclin, 1932; Догель, 1938; Rattenbury, 1954). У последнего, по данным А. О. Ковалевского (1865), жгутиковый эпителий сохраняется вплоть до стадии образования первых жаберных щелей. Давно известно (Hamann, 1885), что у некоторых иглокожих (морские звезды) жгутиковый эпидермис сильно развит и у взрослых особей. Сходство покровного слоя клеток гнатостомулид с таковым бластулообразных личинок Metazoa, а стало быть, и с кино- бластом Фагоцителлы, гипотетического меч- никовского предка многоклеточных животных, имеет глубокий филогенетический смысл. По общему характеру покровов гна- тостомулиды неизмеримо примитивнее, чем большинство взрослых Cnidaria, но сходны с их личинками. Жгутиковый эпидермис мог сохраниться только у форм, стоящих еще близко к общему корню Metazoa; следовательно, наличие его у гнатостомулид говорит против гипотезы происхождения Plat- helminthes от целомических животных. Однако, несмотря на общую значительную архаичность, эпидермис гнатостомулид обладает двумя явно вторичными особенностями: во-первых, он под
стилается базальной мембраной и, во-вторых, лишен эпителиально-мускульных клеток. У гнатостомулид уже полностью закончился процесс обособления кожно-мускульного мешка от эпидермиса — по описанию Акса (Ах, 1956), волокна кожно-мускуль
ного мешка, отличающиеся поперечной исчерченностью, лежат в периферических слоях паренхимы (рис. 38, А). Среди других отрядов турбеллярий сплошной жгутиковый эпидермис неизвестен.
Чрезвычайно примитивное состояние эпидермиса в виде клеточного ресничного эпителия без базальной мембраны, состоящего в основном из эпителиально-мускульных клеток, мы находим
ра
Рис.
9. Oligochoerus
erythrophthalmus (Acoela).
Эпидермис.
(По:
Беклемишев, 1937).
ер
— эпителиально-мускульные клетки, т
— миофибриллы, ра
— клетки паренхимы, г — корешки
ресничек.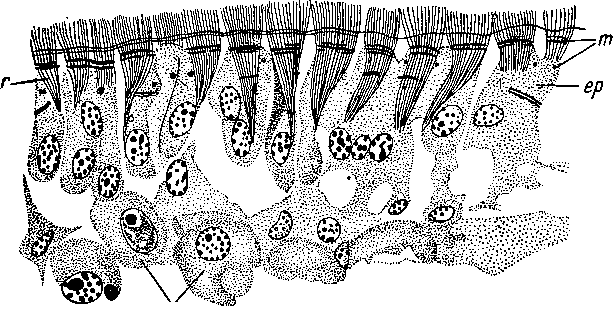
также у Acoela. Такой эпидермис описан В. Н. Беклемишевым (1937) у молодых экземпляров Oligochoerus erythrophthalmus (рис. 9). Он слагается из клеток неправильной формы и различной высоты. Смыкаясь друг с другом, апикальные участки клеток образуют наружную поверхность тела, одетую ресничками. Базальные же концы клеток лежат на разных уровнях и как бы растрепаны, так что в целом весь эпидермис не обособлен от паренхимы и представляет собой нечто вроде ее поверхностного и слабо дифференцированного слоя. В цитоплазме эпидермальных клеток залегают миофибриллы кожно-мускульного мешка — кольцевые и продольные. Из такого примитивного ресничного эпидермиса развиваются другие типы кожного эпителия турбеллярий: погруженный, так называемый немертиновый и обособленный однослойный. Замечательно, что все они встречаются у Acoela.
Существенные эволюционные изменения покровов заключаются, как полагает В. Н. Беклемишев (1925, 1937, 1964), в обособлении сократимого миоэпителиального слоя в виде правильной эпителиальной пластинки, содержащей миофибриллы. Пластинка образу-
ется из смыкающихся апикальных концов клеток, тогда как базальные части клеток с ядрами погружаются в паренхиму. Наиболее примитивная форма погруженного эпидермиса, свойственная большинству Acoela, характеризуется отсутствием базальной мембраны (рис. 10). Однако у представителей других отрядов появляется базальная мембрана, а кожные миофибриллы обособлены от эпидермиса в виде волокон кожно-мускульного мешка, лежащих под эпителиальной пластинкой. Существуют два варианта погруженного эпителия — клеточный и синцитиальный, причем последний образуется независимо в разных группах путем слияния клеточных территорий. Кроме Acoela, погруженный эпидермис имеется у Leciihoepitheliata и у многих
Prolecithophora и Seriata и свойствен Udonellida. Нередко этот,
ер — клетки эпидермиса, gc — железистая клетка, me, md, ml — миофибриллы.
в общем примитивный эпидермис замещается другими, более
Рис.
11. Nemertoderma
bathycola
(Acoela).
Немер-
тиновый эпидермис. (По: Steinbock,
1930—1931).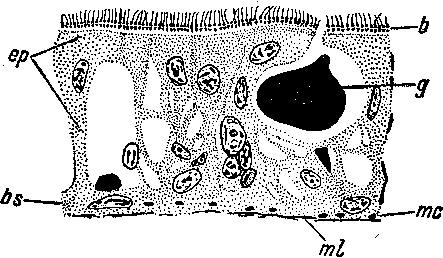
Ь — базальный аппарат ресничек, bs — базальный слой эпидермиса, ер — эпидермальная клетка, g — скопление секрета, тс, ml — миофибриллы.
совершенными типами покровов, сохраняясь лишь на ограниченных участках тела (например, в глотке Baikalarctia gulo, по: Фридман, 1933),йемертиновый эпидермис развит у некоторых Acoela, например у Aechmalotus pyrula и N emertoderma bathycola (рис. 11), и у немногих других очень примитивных турбеллярий. Он очень слабо отграничен от паренхимы и слагается из эпидермальных клеток с узкими базальными концами, между которыми расположены разнообразные клетки как эктодермальной, так и паренхимной природы — железистые, нервные, чувствительные и опорные, соединительнотканные. Немертиновый эпидермис свойствен ближайшим родичам турбеллярий — Xenoturbellida — и встречается также у немертин и у Ente- ropneusta. '
Обособленный однослойный эпителий с хорошо развитой базальной мембраной присущ, как известно, большинству беспозвоночных. Наиболее обычен он и у турбеллярий. Он образуется из примитивного эпидермиса путем правильного, строго однослойного расположения клеток. У Acoela этот тип эпителия встречается довольно часто, но всегда лишен еще базальной мембраны и состоит из эпителиально-мускульных клеток. У других турбеллярий последние дифференцируются на эпидермальные клетки и на мускульные, утратившие связь с эпидермисом и лежащие уже под его базальной мембраной (Беклемишев, 1937, 1964). Обособленный эпителий представляет собой высшую степень отграничения эпидермиса от паренхимы (рис. 12). В некоторых случаях он может принять синцитиальное строение.
Ь — базальный аппарат ресничек, Ьт — базальная мембрана, ер — эпидермальная клетка, d — проток аде- нальной погруженной железистой клетки, g — железистая клетка, г — корешки ресничек, rh — рабдитная железа с рабдитами.
Базальная мембрана
Итак, отсутствие базальной мембраны очень важный признак, особенно подчеркивающий примитивное состояние покровов. Это опорная пластинка, выделяемая, по крайней мере частично, эпителиальными клетками, представляет собой характерный компонент всех эпителиев, и в частности эпидермиса у Metazoa. Она отсутствует у Trichoplax, всех губок и бескишечных турбеллярий. Замечательно, что ее нет также у некоторых низших представителей других отрядов турбеллярий, а именно у двух видов Catenulida — Rhynchoscolex simplex и Rh. diplolithir cus (Reisinger, 1924), у Acanthomacrostomum spiculiferum из Mac-
rostomida (Papi e Swedmark, 1959) и у Hofstenioplesia (=Prorhyn- chus) haswelli из Lecithoepitheliata (Steinbock u. Reisinger, 1924; Bresslau, 1928—1933).
Эпителиально-мускульные клетки
Как мы видели, отсутствие базальной мембраны сочетается с наличием эпителиально-мускульных клеток, весьма напоминающих таковые низших Cnidaria, например гидры. Миофибриллы, принадлежащие эпидермальным клеткам и обычно пронизывающие слой эпидермиса,. были оиисаны у многих Acoela: у Childia
groenlandica и Anaperus tvaermin- nensis (Luther, 1912), Hofstenia atroviridis (Bock, 1923), 'Nemerto- derma bathycola (Steinbock, 1938), Paraphanostoma (Westblad, 1942), Oligochoerus erythrophthalmus, О. ba- kuensis и у Pseudoconvoluta tenera (Беклемишев, 1937, 1960, 1964). Эти миофибриллы слагаются в несколько сплошных слоев — обычно в два слоя — наружный кольцевой и внутренний продольный, которые образуют эпидермальный кожно-мускульный мешок (рис. 13). Последний несомненно гомологичен кожно-мускульному мешку других турбеллярий и представляет собой низшую стадию его эволюционного развития. У представителей других отрядов турбеллярий эпителиально-мускуль- ные клетки встречаются крайне редко и лишь у вышеперечисленных форм, лишенных базальной мембраны.
с — реснички, g — железистая клетка, тс — кольцевые миофибриллы, ml — продольные миофибриллы, п — ядро.
Первичные особенности в строении покровов
Многие зоологи независимо от В. Н. Беклемишева пришли к убеждению, что обе сопряженные друг с другом особенности — наличие эпителиально-мускульных клеток и отсутствие базальной мембраны — относятся к крайне примитивным чертам низших Metazoa (Luther, 1912; Bock, 1923; Bresslau u. Reisinger, 1928—1933; Westblad, 1942; Hyman, 1951; Иванов, 1952a, 1968; Steinbock, 1958; Мамкаев, 1967). Другие авторы считают оба признака вторичными упрощениями, связанными с общей деградацией турбеллярий, и в частности Acoela (Ливанов, 1945; Remane, 1954, 1959; Ах, 1961; Федотов, 1966, и др.).
Акс, например, утверждает, что «методически безупречный» и «точный» анализ отдельных примеров заставляет думать, что эпителиально-мускульные клетки развились как «вторичные струк
турные отношения», а отсутствие базальной мембраны есть результат «вторичного ее исчезновения» (Ах, 1961, стр. 15 и 16). Для катенулид, пишет Акс, характерны базальная мембрана и субэпителиальный кожно-мускульный мешок. Среди видов Rhynchoscolex это нормальное состояние свойственно, например, Rh. evelinae (Marcus, 1945а). Если мы хотим рассматривать эпидермальную мускулатуру и отсутствие базальной мембраны у некоторых видов этого рода как примитивные черты строения, то должны поставить род Rhynchoscolex у корня всех катенулид или считать эти древние признаки очень стойкими. По мнению Акса, против примитивности рода Rhynchoscolex говорят редукция у его представителей мужских половых органов и свойственный им партеногенез. Вторая альтернатива, утверждаем Акс, также неприемлема, потому что в этом случае пришлось бы считать базальную мембрану и типичный кожно-мускульный мешок возникшими у Catenulida по крайней мере дважды.
Аналогичные рассуждения в отношении Acanthomacrostomum убеждают Акса в том, что и здесь имеют место «вторичные структуры». Acanthomacrostomum, — весьма специализированный обитатель песка, который по внутреннему строению, по частичной редукции ресничного покрова и особенно по образованию внутреннего скелета из известковых спикул стоит далеко от основного типа макростомид. Интраэпителиальное положение кожно-мускульного мешка у этой формы, по Аксу, есть результат его вторичного смещения, а отсутствие базальной мембраны — следствие ее редукции.
Аргументы Акса, однако, кажутся убедительными лишь на первый взгляд. Как мы уже подчеркивали, примитивных во всех отношениях организмов в природе не существует. Не представляют исключения, разумеется, и представители Rhynchoscolex. Рейзингер (Reisinger, 1924), детально изучивший организацию Rh. simplex и Rh. diplolithicus, отмечает, что примитивность покровов сочетается у них с многими другими примитивными чертами, отчасти свойственными и другим Catenulida. Так, кишечник у этих видов Rhynchoscolex представлен ацелоидным синцитием, чем отличается от высокодифференцированной эпителизованной кишки у Catenula и Stenostomum. Пищеварение у Rhynchoscolex явно комбинированное — внутриполостное и вместе с тем внутриклеточное. Нервная система отличается примитивными особенностями — у Rhynchoscolex она построена по ортогональному типу, как у Acoela, и обладает по крайней мере четырьмя парами продольных нервных стволов (Reisinger, 1924). С другой стороны, в покровах Rhynchoscoles simplex и Rh. diplolithicus, судя по данным Рейзингера, произошли уже некоторые прогрессивные изменения по сравнению, например, с эпидермисом Oligochoerus erythrophthalmus. У обоих видов в толще эпителия залегают только кольцевые мускульные волокна кожно-мускульного мешка, тогда как продольные уже полностью обособились от эпидермиса и
33
А. В. Иванов и Ю. В. Мамкаев
представлены соответственно миоцитами (или миобластами). Эмансипация кожно-мускульного мешка от эпидермиса остановилась, так сказать, на полпути. В общем, конечно, это состояние покровов у рассматриваемых видов вполне отвечает очень примитивному уровню организации Catenulida.
То
же самое справедливо и в отношении
Acanthomacrostomum
spiculiferum.
С
нашей точки зрения, примитивность его
эпидермиса не противоречит общим
представлениям об эволюции покровов
у низших Metazoa,
развиваемым
В. Н. Беклемишевым. Макростомиды во
многих отношениях несомненно очень
специализированы, но все же находятся
на весьма низком уровне организации,
и проявление у них отдельных архаических
особенностей не представляет ничего
невероятного. Заметим, кроме того, что
Acanthomacrostomum
—
типичный представитель интерстициальной
фауны, в которой, как известно, сохранилось
много удивительно архаических форм,
обладающих в то же время узкими вторичными
адаптациями к весьма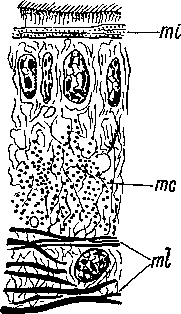
Рис. 14. Hofstenioplesia haswelli (Lecithoepitheliata). Эпидермис и кожно-мускульный мешок. Продольный разрез. (Из: Bresslau, 1928—1933).
me — кольцевые миофибриллы, тг — интразпителиальные миофибриллы, ml — продольные миофибриллы.
специфическим условиям жизни в тесных пространствах между подвижными песчинками.
Наличие эпидермальной мускулатуры у Hofstenioplesia haswelli Акс тоже считает результатом вторичных изменений. По его мнению, об этом свидетельствует принадлежность Hofstenioplesia к высокодифференцированному отряду Lecithoepitheliata. Однако после весьма обстоятельных исследований Штейнбека (Steinbock, 1966) стало совершенно ясно, что Lecithoepitheliata и Hofsteniidae из Acoela обнаруживают целый ряд общих органологических и гистологических особенностей, говорящих о несомненном родстве этих групп (см. также: Reisinger, 1968). В частности, тип строения покровов и кожно-мускульного мешка у Hofsteniidae и у Prorhynchidae один и тот же. У тех и других развит погруженный эпидермис с синцитиальной эпителиальной пластинкой, а кожно-мускульный мешок удвоен, т. е. имеются две системы мускульных слоев — наружная и внутренняя, каждая из кольцевого и продольного слоев (рис. 14). У Hofsteniidae, естественно, нет еще базальной мембраны, а наружные мускульные слои занимают интраэпителиальное положение. То же мы видим и у Hofstenioplesia, наиболее примитивного рода Prorhynchidae, который и по многим другим признакам служит промежуточным звеном между Hofsteniidae и Lecithoepitheliata. У других проринхид — Prorhynchus putealis, Xenoprorhynchus steinbocki — имеется уже базальная мембрана, под которой лежат обе системы кожномускульного мешка (Steinbock, 1927, 1966; Papi, 1957; Reisinger, 1968).
Далее Акс ссылается еще на поучительный, с его точки зрения, случай развития миофибрилл в эпидермисе у одного предста