

Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
118_______________ ГЛАВА 11___________ _______________________________________________________________
|
|
Рис. 11.12. Изолированная экстрахромосомная рРНК из ооцитов Xenopus laevis. На электронных микрофотографиях видны кольца (А) и «лариаты» (структуры типа лассо) (Б), что указывает на репликацию по механизму «катящегося кольца». (Из Rochaix et al., 1974; фотография с любезного разрешения A. Bird.) |
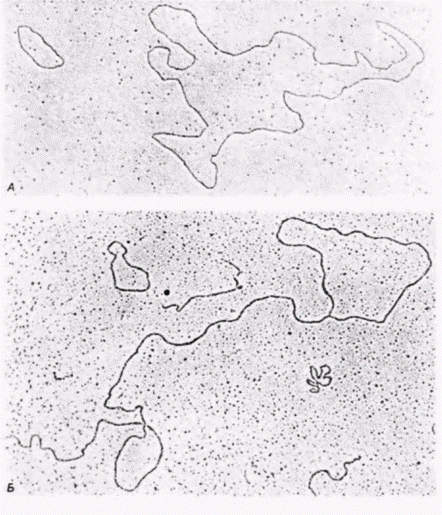
репликации характерен для многих вирусов. В ооците один набор рибосомных генов каким-то образом отсоединяется от хромосомы и сворачивается в кольцо. В кольцо вводится ник (разрыв одной цепи), и с этого ника начинается синтез ДНК в обоих направлениях. Таким путем могут быть синтезированы идентичные копии рибосомных генов и их спейсеров (рис. 11.11). Электронные микрофотографии амплифицированных генов (Hourcade et al., 1973; Rochaix et al., 1974) подтверждают эту модель. На этих микрофотографиях выявляются замкнутые кольца, типичные для экстрахромосомных ядрышек, а также структуры типа «лариатов» (лассо), которые, по-видимому, представляют собой промежуточные продукты катящегося кольца (рис. 11.12).
Визуализация транскрипции на амплифицированных генах
Первыми генами, которые были выделены и очищены, стали амплифицированные гены рибосомных РНК из ооцитов амфибий. Они оказались также первыми генами, транскрипция которых реально наблюдалась с помощью электронного микроскопа. Зная, что эти гены высокоактивны в отношении транскрипции рРНК на стадии диплотены мейоза, Миллер и Битти (Miller, Beatty, 1964) для разделения петель хроматина перед электронной микроскопией использовали буферы с низкой ионной силой, что приводило к расплетанию колец ДНК и возникновению конфигурации, показанной на рис. 11.8. При
Гилберт с. Биология развития: в 3-х т. Т. 2: Пер. С англ. – м.: Мир, 1994. – 235 с.
__________________ ИЗМЕНЕНИЕ ТРАНСКРИПЦИИ В ХОДЕ РАЗВИТИЯ_____________________________________________ 119
этом ДНК образует осевой стержень, на котором синтезируется рРНК. На каждом участке синтезируется множество цепей рРНК. и ряд таких участков показан на этом рисунке. На каждом участке транскрипция начинается на вершине «рождественской елки» и продолжается до тех пор, пока не будет синтезирован предшественник больших рибосомных РНК. В точке присоединения каждого РНК-транскрипта к ДНК находится молекула РНК-полимеразы. Длина РНК в этих больших транскриптах составляет около 7200 оснований, что примерно соответствует ожидаемым размерам предшественника рРНК. Между транскрибируемыми участками ДНК лежат «нетранскрибируемые спейсеры». Электронная микрофотография и в самом деле может дать нам точную картину процесса транскрипции эукариотического гена.
![]()
Дополнительные сведения и гипотезы: Молекулярная основа быстрой транскрипции рибосомных генов
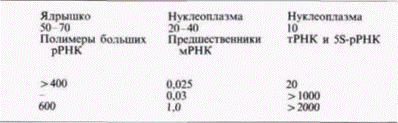
Для осуществления транскрипции необходимо взаимодействие РНК-полимеразы с ДНК. Однако далеко не каждый фрагмент ДНК подходит для этого (в противном случае транскрипция могла бы начаться где угодно и клетка заполнилась бы дефектными молекулами мРНК). Участок ДНК, специализированный для связывания РНК-полимеразы, называют промотором. В клетках эукариот содержатся РНК-полимеразы трех различных типов, каждый из которых характеризуется определенными свойствами и функцией (Rutter et al., 1976). РНК-полимераза I обнаруживается в области ядрышка в ядре и отвечает за транскрипцию больших рРНК, РНК-полимераза II транскрибирует предшественники мРНК, а РНК-полимераза III транскрибирует малые РНК, такие, как транспортные РНК, рибосомную 5S-PHK и другие небольшие последовательности РНК. Эти полимеразы имеют различную чувствительность к ионным условиям и антибиотикам. Их свойства представлены в табл. 11.1.
Высокая эффективность транскрипции рибосомных генов может быть обусловлена структурой области спейсера между соседними генами рибосомной РНК. Как у большинства генов, промотор, присоединяющий РНК-полимеразу, лежит непосредственно перед (5’) кодирующей областью гена. Однако в отличие от большинства других генов у генов рРНК прямо перед промотором имеется последовательность ДНК, отвечающая за терминацию транскрипции (рис. 11.13). Эта структура (до сих пор обнаруженная только в рибосомных генах млекопитающих и амфибий) может концентрировать молекулы РНК-полимеразы на этих генах. У большинства генов при терминации транскрипции происходит высвобождение РНК и РНК-полимеразы в нуклеоплазму. РНК-полимераза может затем использоваться повторно, если найдет на ДНК новый промоторный сайт. Этот процесс, на который заведомо должно тратиться время, возможно, не осуществляется при транскрипции рибосомных генов. Вместо этого после отделения 40S-предшественника рРНК от гена РНК-полимераза остается на спейсере, синтезируя, вероятно, короткие нестабильные транскрипты (Labhart, Reeder, 1986). Как отмечалось ранее, непосредственно перед промотором находится специальный участок терминации транскрипции. Этот участок взаимодействует с промотором для обеспечения переноса РНК-полимеразы к соседнему гену (McStay, Reeder, 1986; Henderson, Sollner-Webb, 1986; Grummt et al., 1986). Поэтому, коль скоро РНК-полимераза начала транскрипцию рибосомного гена, она переносится затем от одного
|
Таблица 11.1. Общая классификация РНК-полимераз эукариот | |||
|
Свойство |
РНК-полимераза I |
РНК-полимераза II |
РНК-полимераза III |
|
|
| ||

![]()