

Нормальная физиология (Пособие для резидентуры)
.pdfНЕЙРОН. КЛАССИФИКАЦИЯ НЕЙРОНОВ |
5 |
|
|
жен. Если на нерв наносится несколько слабых раздражений, т.е. торможение оказывается усиленным, то наблюдается суммация тормозных влияний.
И.П.Павлов показал, что явление торможения имеет важное значение во всех проявлениях высшей нервной деятельности и поведения организма.
Торможение в след за возбуждением. Особым видом торможения является торможение, развивающееся в нервной клетке после прекращения ее возбуждения. Оно возникает в том случае, если после окончания вспышки возбуждения, в клетке развивается сильная следовая гиперполяризация мембраны. Возбуждающий постсинаптический потенциал в этих условиях оказывается недостаточным для критической деполяризации мембраны, и распространяющееся возбуждение не возникает.
5.5. Координациярефлекторныхпроцессов
Взаимодействие нейронов, а следовательно, и нервных процессов в центральной нервной системе, обеспечивающее ее согласованную деятельность, носит название координации. Координация обеспечивает точное выполнение мышечных движений, создает приспособленные к различным внешним ситуациям рефлекторные акты, в состав которых входят двигательные, сосудистые, секреторные и другие компоненты.
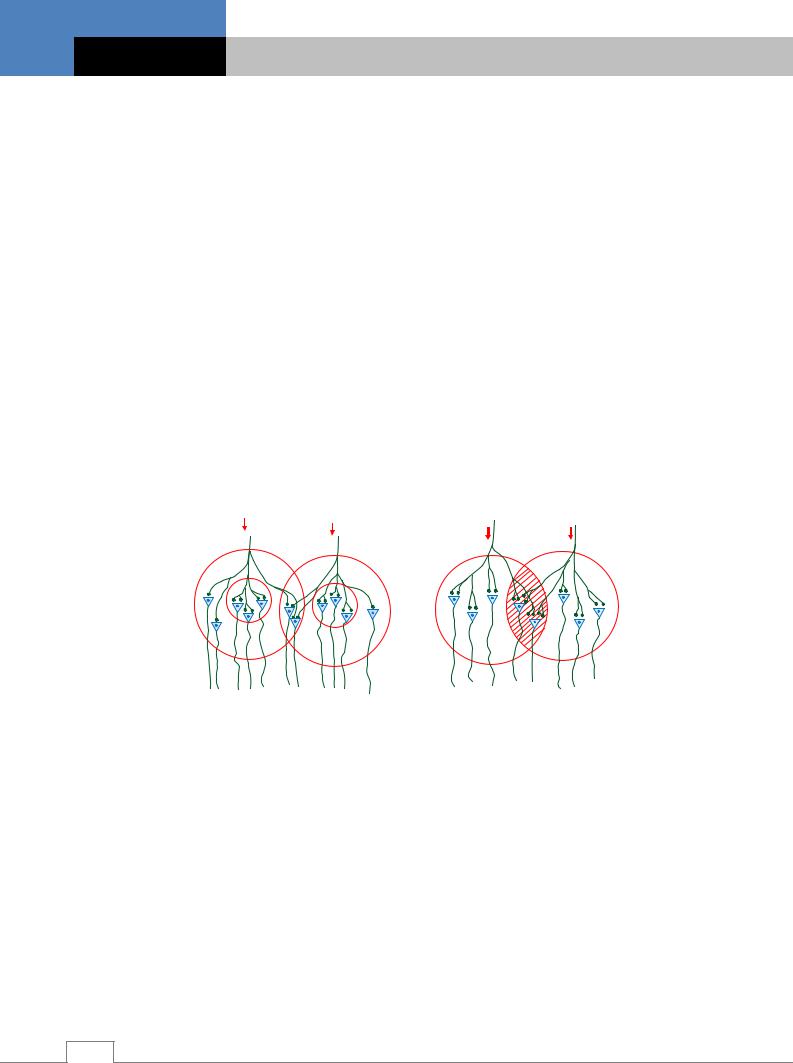
Конвергенция. Импульсы, приходящие в центральную нервную систему по различным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же промежуточным и эффекторным нейронам. Этот факт лег в основу принципа конвергенции установленного Ч.Шеррингтоном. Конвергенция нервных импульсов объясняется тем, что на теле и дендритах каждого нейрона центральной нервной системы оканчиваются аксоны множества других нервных клеток. Конвергируют афферентные импульсы, возникающие в различных участках рецептивного поля только одного и того же рефлекса. В отличие, от этого в высших отделах центральной нервной системы, в подкорковых ядрах и в больших полушариях наблюдается конвергенция импульсов, исходящих из разных рецептивных зон. Поэтому один и тот же нейрон может возбуждаться импульсами, возникающими при раздражении и слуховых, и зрительных, и кожных рецепторов (мультисенсорная конвергенция). Другой вид конвергенции мультибиологический – заключается в схождении к одному нейрону возбуждений от центров, формирующих те или иные биологические состояния (голод, жажда). Третий вид конвергенции – сенсорно-биологический – представляет собой комбинацию двух первых, например, схождение к нейрону импульсов от рецепторов сетчатки глаза и гипоталамического центра голода (рис. 5.21) /4/.
|
|
|
Рис. 5.21. Динамика процесса конвергенции. |
|
|
|
Схождение импульсов, идущих по множеству |
|
|
|
афферентных волокон к одному нейрону: |
|
|
|
А – мультисенсорная конвергенция; |
|
|
|
Б – мультибиологическая конвергенция; |
Свет |
Звук Голод |
Жажда Свет Голод |
В – сенсорно-биологическая конвергенция. |
|
А Б В
101
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Конвергенция объясняет пространственную суммацию возбуждений и явление окклюзии. Нервные центры состоят из большого числа нейронных популяций, причем отдельные нейроны могут входить в разные популяции. Каждый аксон рецепторного нейрона, поступая в центральную нервную систему, дает большее количество разветвлений и образует синапсы на целой популяции нервных клеток – нейронном «пуле». В такой популяции принято условно различать центральную-пороговую зону и периферическую-подпороговую «кайму». Облегчение. При раздельном раздражении афферентных волокон I и II (рис. 5.22А) возникают рефлекторные реакции, сила которых при прочих равных условиях определяется только числом возбужденных нейронов центральной пороговой зоны. При совместном же раздражении этих афферентных волокон возбуждающие постсинаптические потенциалы, возникающие в нейронах подпороговой «каймы» суммируются друг с другом и достигают критической величины. В результате в процесс возбуждения оказываются вовлеченными нейроны не только центральной зоны (6 нейронов), но и нейроны подпороговой «каймы» (2), на которых конвергируют разветвления этих волокон. Поэтому сила рефлекторной реакции оказывается большей, чем арифметическая сумма реакций, вызываемых раздельными раздражениями волокон I и II (6+2 6). Такое явление получило название центрального облегчения (рис. 5.22А). Оно чаще всего наблюдается при слабых раздражениях, при которых акти-
вируется лишь небольшое число афферентных нервных волокон.
I |
II |
I |
II |
А |
Б |
Рис. 5.22. Схема, иллюстрирующая явления облегчения (А) и окклюзии (Б).
Окклюзия. Отдельные нейроны могут входить в центральные зоны разных нейронных популяций. В таких случаях число нейронов, возбуждаемых при совместном раздражении афферентных волокон I и II, 8 оказывается меньше арифметической суммы числа нейронов, возбуждаемых при раздельном раздражении этих волокон (8 10). Это явление было названо Шеррингтоном явлением окклюзии (закупорки) (рис. 5.22Б).
При наличии окклюзии суммация возбуждающих постсинаптических потенциалов на телах нейронов, находящихся в области совпадения центральных зон, не приводит к усилению рефлекторной реакции, поскольку возбуждающие постсинаптические потенциалы, вызываемые каждым из этих раздражений в отдельности, имеют пороговую величину. Поэтому суммарный эффект совместного раздражения I и II будет меньше арифметической суммы этих эффектов, полученных порознь. Явление окклюзии хорошо выражено в случае применения
102
НЕЙРОН. КЛАССИФИКАЦИЯ НЕЙРОНОВ |
5 |
|
|
сильных афферентных раздражений, каждое из которых активирует большое число нейронов.
Сущность окклюзии (закупорки) состоит в том, что при одновременном раздражении двух групп афферентных волокон, каждая из которых вызывает сильные рефлекторные сокращения мышцы возникает эффект, который по своей величине оказывается меньше арифметической суммы величин этих рефлексов, взятых порознь. На рис. 5.22Б представлены моторные нейроны, получающие импульсы от двух групп афферентных волокон. При максимальном раздражении каждой из этих групп в отдельности, возбуждение охватывает по 5 нейронов. При совместном же раздражении волокон I и II возбуждение возникает не в 10, а только в 8 нейронах, так как две нервные клетки вследствие конвергенции иннервируются обеими группами нервных волокон. Поэтому рефлекторный ответ совместного раздражения волокон I и II меньше арифметической суммы этих рефлексов, полученных порознь. Это явление объясняется частичным перекрытием рецепторных синаптических полей двух рефлекторных реакций (рис. 5.22Б).
В нормальной деятельности нервных центров пространственная суммация и окклюзия постоянно совмещаются друг с другом, становясь то более, то менее выраженными, в зависимости от силы взаимодействующих афферентных раздражений: при слабой их силе на первый план выступает суммация, а при сильных раздражениях – окклюзия.
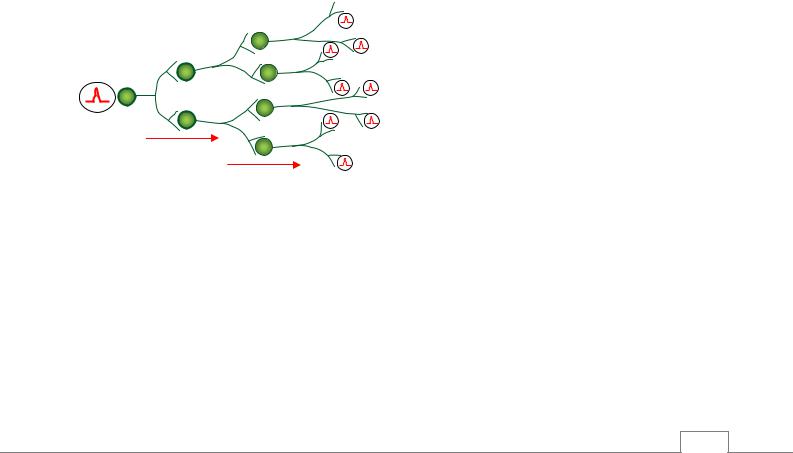
Иррадиация возбуждения. Импульсы, поступающие в ЦНС при сильном и длительном раздражении, вызывают возбуждение не только нейронов данного рефлекторного центра, но и других нервных центров. Это распространение возбуждения в ЦНС получило название ир-
радиации (рис. 5.23).
Рис. 5.23. Механизм иррадиации. Показано распространение импульсов, поступающих по афферентным волокнам от одного нейрона через коллатерали его аксона к большему числу нервных клеток /4/.
Для иллюстрации приведем следующий опыт. Если у кошки после перерезки мозгового ствола, отделяющего вышележащие отделы головного мозга (например, большие полушария) от нижележащих, нанести слабые раздражения на подошву задней лапы, то наблюдается сгибание лапы только в голеностопном суставе. Усиление раздражения приведет к сгибанию, кроме того, еще и в коленном суставе, а еще большее усиление – к сгибанию в тазобедренном суставе.
Иррадиация может быть системной (направленной), например, при вытягивании руки возбуждается только нейроны экстензоров, а флексорные нейроны тормозятся, и диффузной, например, при введении стрихнина, блокирующего тормозные синапсы.
103

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Иррадиация обусловлена многочисленными ветвлениями отростков (аксонов и дендритов) нервных клеток и цепями вставочных нейронов, объединяющих друг с другом различные нервные центры. Особую роль в механизме иррадиации возбуждения играет ретикулярная формация. Иррадиации возбуждения препятствуют многочисленные тормозные нейроны и синапсы, входящие в состав различных рефлекторных центров.
Последовательная смена возбуждения торможением и торможения возбуждением.
При исследовании механизмов координации рефлекторных актов часто встречаются контрастные изменения состояния нервного центра, возникающие после прекращения возбуждающего или тормозящего раздражения. Эти изменения состоят в том, что вслед за торможением наблюдается усиление возбуждения («положительная последовательная индукция»), а вслед за возбуждением – торможение («отрицательная последовательная индукция»).
Возникновение вспышки возбуждения в нервном центре после выключения тормозящего раздражителя впервые было описано И.М.Сеченовым. Позднее Шеррингтон наблюдал, что если раздражать кожу на боку собаки, что вызывает слабый рефлекс почесывания, и присоединять электрическое раздражение кожи лапы, происходит торможение чесательного рефлекса, который, резко усиливается после прекращения тормозящего раздражения. Примером торможения вслед за возбуждением может служить следующее, описанное А.А.Ухтомским явление. К ритмическому раздражению нерва, вызывающему рефлекс сгибания лапки лягушки, присоединяют раздражение другого нерва, вызывающего тот же рефлекс сгибания. Прекращение второго раздражения сопровождается резким уменьшением рефлекса по сравнению с тем, что наблюдался вначале.
Явления индукции могут наблюдаться в деятельности всех отделов центральной нервной системы. И.П.Павловым открыты явления индукции, происходящие в коре больших полушарий головного мозга. Контрастное усиление возбуждения вслед за торможением и торможение вслед за возбуждением, И.П.Павлов наблюдал при изучении условно-рефлекторной деятельности и назвал их соответственно явлением последовательной корковой положительной или отрицательной индукции.
104

ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ |
6 |
|
|
ГЛАВА6. ЧАСТНАЯФИЗИОЛОГИЯЦЕНТРАЛЬНОЙ НЕРВНОЙСИСТЕМЫ
6.1. Спинноймозг
Спинной мозг человека имеет 31-33 сегментов: 8 шейных (СI-СVIII), 12 грудных (ThI-ThXII), 5 поясничных (LI-LV), 5 крестцовых (SI-SV), 1-3 копчиковых (CоI-CоIII) /3/.
Морфологических границ между сегментами спинного мозга не существует, поэтому деление на сегменты является функциональным и определяется зоной распределения в нем волокон заднего корешка и зоной клеток, которые образуют выход передних корешков.
В опытах с перерезкой и раздражением корешков спинного мозга показано, что задние корешки являются афферентными (чувствительными), а передние эфферентными (двигательными), закон Белла-Мажанди /3/.
Афферентные входы в спинной мозг организованы аксонами спинальных ганглиев, лежащих вне спинного мозга, и аксонами экстра- и интрамуральных ганглиев симпатического и парасимпатического отделов автономной нервной системы.
Первая группа афферентных входов спинного мозга образована чувствительными волокнами, идущими от мышечных рецепторов, рецепторов сухожилий, надкостницы, оболочек суставов. Эта группа рецепторов образует начало проприорецептивной чувствительности.
Вторая группа афферентных входов спинного мозга начинается от кожных рецепторов: болевых, температурных, тактильных, давления – и представляет собой кожную рецептирующую систему.
Третья группа афферентных входов спинного мозга представлена рецептирующими входами от висцеральных органов, это висцерорецептивная система.
Эфферентные (двигательные) нейроны расположены в передних рогах спинного мозга, их волокна иннервируют все скелетные мышцы /3/.
Особенностинейроннойорганизацииспинногомозга
Спинной мозг человека содержит около 13 млн. нейронов, из них 3% мотонейроны, а 97% вставочные /3/. Функционально нейроны спинного мозга можно разделить на 4 основные группы:
 мотонейроны, или двигательные клетки передних рогов, аксоны которых образуют передние корешки;
мотонейроны, или двигательные клетки передних рогов, аксоны которых образуют передние корешки;  интернейроны – нейроны, получающие информацию от спинальных ганглиев и рас-
интернейроны – нейроны, получающие информацию от спинальных ганглиев и рас-
полагающиеся в задних рогах, реагируют на болевые, температурные, тактильные, вибрационные, проприорецептивные раздражения;
105

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
 симпатические и парасимпатические нейроны расположены преимущественно в боковых рогах, аксоны этих нейронов выходят из спинного мозга в составе передних корешков;
симпатические и парасимпатические нейроны расположены преимущественно в боковых рогах, аксоны этих нейронов выходят из спинного мозга в составе передних корешков;
 ассоциативные клетки – нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами /3/.
ассоциативные клетки – нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами /3/.
Мотонейроны. Аксон мотонейрона своими окончаниями снабжает мышечные волокна, образуя мотонейронную единицу. Чем меньше мышечных волокон иннервирует один аксон
(т.е. чем меньше количественно мотонейронная единица) тем более дифференцированные, точные движения выполняет мышца /3/. Несколько мотонейронов могут иннервировать одну мышцу – в этом смысле они образуют так называемый мотонейронный пул (моторное поле) (рис. 6.1).
Мотонейроны спинного мозга функционально делят на α- и γ-мотонейроны. α-Мотонейроны иннервируют скелетную мускулатуру (экстрафузальные волокна), обеспечивая мышечные сокращения. γ-Мотонейроны иннервируют рецепторы растяжения (интрафузальные волокна). Благодаря сочетанной активации α- и γ-мотонейронов рецепторы растяжения могут активироваться не только во время растяжения мышц, но и при их сокращении, что важно для обеспечения моторной координации /3/.
Рис. 6.1. Моторное поле мышцы /4/.
Интернейроны, или промежуточные нейроны генерируют импульсы с частотой более 1000 в сек. Функция интернейронов заключается в организации связей между структурами спинного мозга и обеспечении влияния восходящих и нисходящих путей на клетки отдельных сегментов спинного мозга. К числу промежуточных нейронов относятся клетки Реншоу. Возбуждение интернейронов, связанных с моторными клетками, оказывает тормозящее влияние на мышцы – антагонисты /3/.
Фракционирование моторного поля. Каждое афферентное волокно обслуживает только
часть моторного поля (рис. 6.2). На раздражение чувствительного нерва мышца поднимает меньший груз, чем на раздражение двигательного нерва. Каждый чувствительный нерв возбуждает только часть моторного поля мышцы /4/.
Возбудимость мотонейронов одного пула различна, поэтому при разной интенсивности раздражения в сокращение вовлекается разное количество волокон одной мышцы. При оптимальной силе раздражения сокращаются все волокна данной мышцы, в этом случае развивается максимальное сокращение мышцы /3/.
Рис. 6.2. Фракционирование моторного поля для отдельных мышечных волокон /4/.
Проводящие пути спинного мозга (табл. 6.1). Белое вещество спинного мозга состоит из миелиновых волокон, которые могут быть короткими (межсегментарные) и длинными (соединяющими разные отделы головного мозга со спинным и наоборот) /3/.
106

ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ |
6 |
|
|
Короткие волокна (ассоциативные) связывают нейроны разных сегментов или симметричные нейроны противоположных сторон спинного мозга.
Таблица6.1. Основныепроводящиепутиспинногомозга
Проводящиепути |
|
Столбыспинного |
Физиологическоезначение |
|
|
мозга |
|
|
|
|
|
А. восходящие(чувствительные) пути |
|||
Тонкий пучок (пучок Голля) |
|
Задние |
Тактильная чувствительность, чувства |
|
|
|
положения тела, пассивных движений тела, |
|
|
|
вибрации |
|
|
|
|
Клиновидный пучок |
|
Задние |
Тактильная чувствительность, чувства |
(пучок Бурдаха) |
|
|
положения тела, пассивных движений тела, |
|
|
|
|
|
|
|
вибрации |
|
|
|
|
Дорсолатеральный |
|
Боковые |
Пути болевой и температурной |
|
|
|
чувствительности |
|
|
|
|
Дорсальный спиномозжечковый |
|
Боковые |
Импульсы из проприорецепторов мышц, |
Флексига |
|
|
сухожилий, связок; чувство давления и |
|
|
|
прикосновения из кожи |
|
|
|
|
Вентральный спиномозжечковый |
|
Боковые |
Импульсы из проприорецепторов мышц, |
(Говерса) |
|
|
сухожилий, связок; чувство давления и |
|
|
|
прикосновения из кожи |
|
|
|
|
Дорсальный спиноталамический |
|
Боковые |
Болевая и температурная чувствительность |
|
|
|
|
Спинотектальный |
|
Боковые |
Сенсорные пути зрительно-двигательных |
|
|
|
рефлексов и болевой чувствительности |
|
|
|
|
Вентральный |
|
Передние |
Тактильная чувствительность |
спиноталамический |
|
|
|
|
|
|
|
|
Б. Нисходящие(двигательные) пути |
||
Латеральный кортико-спиналь- |
|
Боковые |
Импульсы к скелетным мышцам. |
ный (пирамидный) |
|
|
Произвольные движения |
|
|
|
|
Руброспинальный (Монакова) |
|
Боковые |
Импульсы, поддерживающие тонус |
|
|
|
скелетных мышц |
|
|
|
|
Дорсальный |
|
Боковые |
Импульсы, обеспечивающие поддерживание |
вестибулоспинальный |
|
|
позы и равновесия тела |
|
|
|
|
Оливоспинальный (Гельвега) |
|
Боковые |
Функция неизвестна. Возможно, он |
|
|
|
участвует в осуществлении |
|
|
|
таламоспинальных рефлексов |
|
|
|
|
Ретикулоспинальный |
|
Передние |
Импульсы поддерживающие тонус |
|
|
|
скелетных мышц, регулирующие состояние |
|
|
|
спинальных вегетативных центров и |
|
|
|
чувствительность мышечных веретен |
|
|
|
проприорецепторов скелетных мышц |
|
|
|
|
Вентральный |
|
Передние |
Импульсы, обеспечивающие поддержание |
вестибулоспинальный |
|
|
позы и равновесия тела |
|
|
|
|
Тектоспинальный |
|
Передние |
Импульсы, обеспечивающие осуществление |
|
|
|
зрительных и слуховых двигательных |
|
|
|
рефлексов (рефлексов четверохолмия) |
|
|
|
|
Вентральный кортико-спиналь- |
|
Передние |
Импульсы к скелетным мышцам, |
ный (пирамидный) |
|
|
произвольные движения |
|
|
|
|
107

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Длинные волокна (проекционные) делятся на восходящие, идущие к головному мозгу, и нисходящие – идущие от головного мозга к спинному. Эти волокна образуют проводящие пути спинного мозга. Пучки аксонов образуют вокруг серого вещества так называемые канатики: передние – расположенные кнутри от передних рогов, задние – расположенные между задними рогами серого вещества, и боковые – расположенные на латеральной стороне спинного мозга между передними и задними корешками.
Впередних канатиках расположены нисходящие пути /3/:
передний корково-спинномозговой, или пирамидный путь (tractus corticospinalis ventralis, s.anterior), являющийся прямым неперекрешенным;
покрышечно-спинномозговой, или тектоспинальный путь (tractus teсtospinalis);
преддверно-спинномозговой, или вестибулоспинальный путь (tractus vestibulospinalis).
Взадних канатиках проходят восходящие пути /3/:
тонкий пучок, или пучок Голля (fasciculus gracilis);
клиновидный пучок, или пучок Бурдаха (fasciculus cuneatus).
В боковых канатиках проходят нисходящие и восходящие пути. К нисходящим путям относятся:
 латеральный корково-спинномозговой, или пирамидный, путь (tractus corticospinalis lateralis), является перекрещенным;
латеральный корково-спинномозговой, или пирамидный, путь (tractus corticospinalis lateralis), является перекрещенным;
 красноядерно-спинномозговой, или руброспинальный, путь (tractus rubrospinalis);
красноядерно-спинномозговой, или руброспинальный, путь (tractus rubrospinalis);
 ретикуло-спинальный путь (tractus reticulospinalis).
ретикуло-спинальный путь (tractus reticulospinalis).
К восходящим путям относятся:
 спиноталамический путь (tractus spinotalamicus);
спиноталамический путь (tractus spinotalamicus);
 латеральный и передний спинно-мозжечковые пути, или пучки Флексига и Говерса
латеральный и передний спинно-мозжечковые пути, или пучки Флексига и Говерса
(tractus spinocerebellaris lateralis et ventralis).
Ассоциативные, или проприоспинальные, пути связывают между собой нейроны одного или разных сегментов спинного мозга. Их функция заключается в координации позы, тонуса мышц, движений разных метамеров туловища. К проприоспинальным путям относятся также комиссуральные волокна, соединяющие функционально однородные симметричные и несимметричные участки спинного мозга /3/.
Рефлекторныефункцииспинногомозга
Собственная рефлекторная деятельность спинного мозга осуществляется сегментарными рефлекторными дугами.
Сегментарная рефлекторная дуга состоит из рецептивного поля, от которого, импульсация по чувствительному волокну нейрона спинального ганглия, а затем по аксону этого же нейрона через задний корешок входит в спинной мозг. Далее аксон может идти прямо к мотонейрону переднего рога, аксон которого подходит к мышце. Так образуется моносинаптическая рефлекторная дуга, которая имеет один синапс между афферентным нейроном спинального ганглия и мотонейроном переднего рога. Эти рефлекторные дуги образуются в таких рефлексах, которые возникают только при раздражении рецепторов аннулоспиральных окончаний мышечных веретен.
108
ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ |
6 |
|
|
Другие спинальные рефлексы реализуются с участием интернейронов заднего рога или промежуточной области спинного мозга. В итоге возникают полисинаптические рефлекторные дуги (рис. 6.3).
А |
Б |
Рис. 6.3. |
Проведение |
возбуждений |
по |
моносинаптической и полисинаптической |
|||||
|
|
рефлекторным дугам: А – возбуждение |
|||
|
|
рецепторов |
мышечного |
веретена |
и |
Рпроведение возбуждения по афферентному и
|
|
эфферентному |
|
нейронам |
в |
|
|
|
моносинаптической дуге; |
|
|
||
|
|
Б – возбуждение внешним раздражителем |
||||
МВ |
|
рецептора; проведение возбуждения |
по |
|||
|
афферентному |
нейрону, |
нескольким |
|||
|
|
|||||
|
|
вставочным и эфферентному нейрону в |
||||
|
|
полисинаптической |
дуге. |
Отмечается |
||
|
|
большее время рефлекса. |
|
|
||
Р |
Р |
Р – раздражения /4/. |
|
|
|
|
|
|
|
|
|
||
Рассмотрим некоторые рефлексы спинного мозга.
Миотатические рефлексы – рефлексы на растяжение мышцы. Быстрое растяжение мышцы всего на несколько миллиметров механическим ударом по ее сухожилию приводит к сокращению всей мышцы и двигательной реакции. Например, легкий удар по сухожилию надколенной чашечки вызывает сокращение мышцы бедра и разгибание голени.
Висцеромоторные рефлексы возникают при стимуляции афферентных нервов внутренних органов и характеризуются появлением двигательных реакций мышц грудной клетки и брюшной стенки, мышц разгибателей спины.
Рефлексы автономной (вегетативной) нервной системы обеспечивают реакцию внутренних органов сосудистой системы на раздражение висцеральных, мышечных, кожных рецепторов.
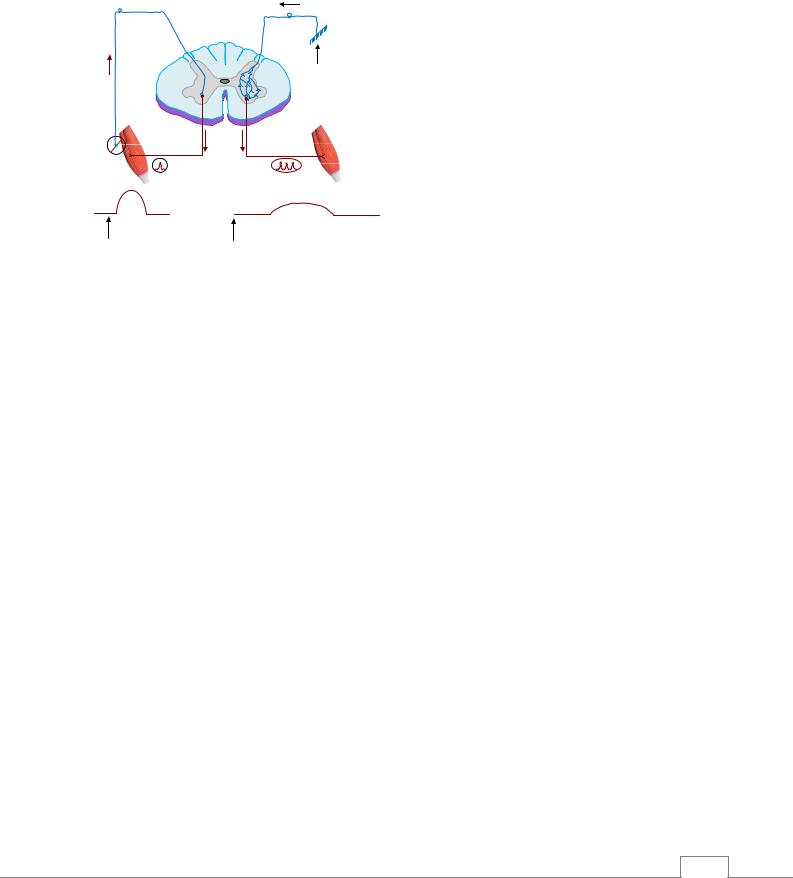
При травмах у человека в ряде случаев происходит полное или половинное пересечение спинного мозга. При половинном повреждении спинного мозга развивается синдром БроунСекара (рис. 6.4). Он проявляется в том, что на стороне поражения спинного мозга (ниже места поражения) развивается паралич двигательной системы вследствие повреждения пирамидных путей, а также нарушается проприорецептивная чувствительность. Это обусловлено тем, что восходящие пути глубокой чувствительности идут по своей стороне спинного мозга до продолговатого мозга, где происходит их перекрест. На противоположной поражению стороне движения сохраняются, отмечается потеря болевой и температурной чувствительности. В итоге, если повреждена левая половина спинного мозга, исчезает болевая чувствительность правой половины туловища ниже повреждения /3/.
Ошушение легкого прикосновения на стороне поражения ослабляется, поскольку пересекается главный путь его передачи – задние столбы. Известно, что волокна в составе этих столбов не переходят на противоположную сторону вплоть до достижения ими продолговатого мозга. Ощущения грубого прикосновения, нечетко локализованного, еще существует, поскольку частично оно передается по спиноталамическому тракту противоположной стороны.
109

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Сторона перерезки Интактная сторона
|
|
Рис. 6.4. Синдром Броун-Секара. |
|
|
Нарушение болевой и |
Зоны поражения у человека при |
|
Нарушение движения |
возбуждении спинного мозга на |
||
температурной чувствительности |
|||
|
уровне грудных сегментов. |
||
|
Частичное поражение тактильной и |
|
|
|
проприоцептивной чувствительности |
|
Спинальный шок. Наблюдается после полной поперечной перерезки спинного мозга. При этом все функции спинного мозга исчезают. У лягушек рефлекторные реакции спинного мозга быстро (через 10-15 мин.) восстанавливаются. У человека восстановление функций разорванного спинного мозга практически не происходит /1/.
Микроэлектродные исследования показали, что при спинальном шоке мотонейроны не страдают. При этом повреждаются вставочные нейроны вследствие чего отсутствуют реакции на афферентные воздействия.
Животные с перерезкой между спинным и продолговатым мозгом получили название спинальных животных. У спинальных животных отсутствует самостоятельное дыхание, наблюдается низкое кровяное давление; снижен сосудистый тонус; утрачена способность поддерживать постоянство температуры тела; исчезают все формы сложной целенаправленной деятельности (пищевая, половая, оборонительная). При перерезках ниже люмбальных отделов наблюдается паралич внешних анальных и мочеиспускательных сфинктеров /4/.
У спинальных животных различают два вида рефлексов на растяжение: фазические и тонические рефлексы. При кратковременном растяжении мышцы, вызванном ударом по сухожилию, возникает фазический сухожильный рефлекс (коленный, ахиллов) и др. Длительное растяжение мышцы сопровождается ее продолжительным сокращением, и эта рефлекторная реакция называется тоническим рефлексом. Причина возникновения этих рефлексов общая – возбуждение рецепторов мышечных веретен растягиваемой мышцы, а эффекторы разные. Считают, что при фазических сухожильных рефлексах преимущественно возбуждаются быстрые мышечные волокна, а в тонических рефлексах участвуют медленные α-мотонейроны и медленные красные мышечные волокна.
Вегетативные рефлексы спинного мозга /1, 4/:
 дефекация;
дефекация;
 мочеиспускание;
мочеиспускание;
 регуляция сосудистого тонуса;
регуляция сосудистого тонуса;
 эрекция и эякуляция;
эрекция и эякуляция;
 потоотделение.
потоотделение.
Рефлексы, которые обычно исследуют у человека, приведены в табл. 6.2.
110