

Нормальная физиология (Пособие для резидентуры)
.pdfРегистрация суммарного
потенциала действия
Раздражение
Проведение
возбуждения
α βγ δ
А В С 
Нерв
ФИЗИОЛОГИЯ НЕРВНЫХ ВОЛОКОН. СИНАПСЫ |
4 |
|
|
Рис. 4.7. Изменение формы суммарного потенциала действия нерва за счет разной скорости проведения возбуждения по разным группам нервных волокон:
Аα, Аβ, Аγ, Аδ, В, С – группы нервных волокон /5/.
Три другие группы волокон типа А: Аβ, Аγ и Аδ – имеют меньший диаметр, меньшую скорость проведения возбуждения и более длительный потенциал действия. Это преимущественно чувствительные волокна, проводящие возбуждение от различных рецепторов в ЦНС. Волокна Аβ образуют афферентные волокна от рецепторов прикосновения кожи в ЦНС. Волокна Аγ – образуют эфферентные волокна к мышечным веретенам. Волокна Аδ являются преимущественно афферентными соматическими волокнами, идущими от температурных и болевых рецепторов кожи в ЦНС. При проведении возбуждения по этим волокнам возникает первичный электрический компонент боли.
Волокна типа В – тонкие миелинизированные, диаметром 10-11 мкм и скоростью проведения возбуждения менее 10 м/сек. Продолжительность потенциала действия волокон типа В (1-2 мс) примерно в 3 раза превышает длительность потенциала действия волокон типа А. Отличительной особенностью этих волокон является то, что в них отсутствует фаза следовой деполяризации; нисходящее колено пика непосредственно переходит в следовую гиперполяризацию, которая в ряде случаев продолжается более 100 мс. Входят в состав преганглионарных волокон вегетативной нервной системы.
Волокна типа С – тонкие безмиелиновые, диаметром 4-5 мкм, скоростью проведения возбуждения 0,7 м/сек. Потенциалы действия волокон этого типа характеризуются наибольшей продолжительностью (2 мс у теплокровных). Входят в состав постганглионарных волокон вегетативной нервной системы. Волокна типа С также передают афферентное возбуждение от внутренних органов в ЦНС. По ним передается возбуждение, формирующее вторичный протопатический компонент боли.
Характеристика проведения возбуждения в смешанном нерве представлена на табл. 4.1.
61

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Таблица 4.1.Проведение возбуждения в смешанном нерве /6/.
Тип |
Диаметр |
Скорость |
Длительность |
Характерпередаваемойинформации |
|
волокна |
волокна, |
проведения, |
ПД, мс |
|
|
|
мкм |
м/с |
|
|
|
|
|
100-160, |
|
Произвольные двигательные волокна, |
|
Аα |
12-20 |
0,4-05 |
проприорецепция, мышечная чувстви- |
||
(70-120) |
|||||
|
|
|
тельность |
||
|
|
|
|
||
|
|
|
|
Осязание, прикосновение (тактильная |
|
Аβ |
8-12 |
40-70 |
0,4-0,6 |
||
чувствительность), давление |
|||||
|
|
|
|
||
|
|
|
|
Чувствительные афферентные волокна от |
|
|
|
|
|
||
А |
4-8 |
15-40 |
0,5-0,7 |
задних корешков спинного мозга, |
|
эфферентные волокна к мышечным |
|||||
|
|
|
|
||
|
|
|
|
веретенам |
|
|
|
|
|
Проведение острой болевой, температур- |
|
А |
1-4 |
5-15 |
0,6-1,0 |
||
ной, тактильной чувствительности |
|||||
|
|
|
|
||
|
|
|
|
|
|
В |
10-11 |
10 |
1-2 |
Преганглионарные волокна ВНС |
|
|
|
|
|
Боль, температура, некоторые виды меха- |
|
|
|
|
|
||
С |
4-5 |
0,7-1 |
2,0 |
норецепции, постганглионарные волокна |
|
|
|
|
|
ВНС |
|
|
|
|
|
|
4.4. Физиологиясинапсов
Синапс – это специализированная структура, которая обеспечивает передачу возбуждения от одной возбудимой структуры к другой. Термин «синапс» был введен в
физиологию Ч.Шеррингтоном, что означает «соединение», «застежка» /4/. Синапс представляет собой сложную структуру и состоит из пресинаптической части (окончание аксона, передающее сигнал), синаптической щели и постсинаптической части (структура воспринимающей клетки) (рис. 4.8).
|
Безмиелиновое окончание |
|
двигательного нерва |
|
Пузырьки с медиатором |
Синаптическая |
Пресинаптическая мембрана |
|
Эндоплазматическая сеть |
Митохондрия |
|
 Са2+ Са2+
Са2+ Са2+
Постсинаптическая мембрана с хеморецепторами
Рис. 4.8. Ультраструктура синапса (схема) /4, 6/.
62

ФИЗИОЛОГИЯ НЕРВНЫХ ВОЛОКОН. СИНАПСЫ |
4 |
|
|
Пресинаптическое окончание аксона нейрона при подходе к иннервируемой клетке теряет миелиновую оболочку, что несколько снижает скорость распространения волны возбуждения. Небольшое утолщение на конце волокна, называемое синаптической бляшкой, содержит синаптические пузырьки с медиатором – веществом, способствующим передаче возбуждения в синапсе. В пресинаптической области обнаружены митохондрии, гранулы гликогена, спиралевидные нити – филаменты.
Синаптическая щель – пространство между пресинаптическим окончанием и участком мембраны эффекторной клетки является непосредственным продолжением межклеточного пространства; ее содержимое – гель, в состав которого входят гликозаминогликаны /6/.
Постсинаптическая мембрана – участок эффекторной клетки, контактирующий с пресинаптической мембраной через синаптическую щель. От постсинаптической мембраны по направлению к ядру клетки прослеживаются нежные микротрубочки, образованные молекулами специфических белков. Полагают, что им принадлежит определенная роль в распространении и обработке информации внутри клетки. Уникальной структурой постсинаптической мембраны являются клеточные рецепторы – сложные белковые молекулы, способные к конформации, т.е. изменяющие пространственную ориентацию при взаимодействии с соответствующими им веществами – лигандами /6/. В результате конформации в центрах связывания рецептора с медиатором изменяется проницаемость мембранных ионоселективных каналов эффекторной клетки. Это в свою очередь способствует возбуждению или торможению эффекторной клетки.
Организации синапсов способствуют специальные белковые вещества, которые вырабатываются в нервной ткани и называются синаптинами. Кроме того, организации синапсов способствуют также факторы роста нервов и инсулин. Если синапс структурно создается нервом, то его физиологические свойства зависят от свойств постсинаптической мембраны иннервируемой ткани.
Исключительную роль синапсов в процессах жизнедеятельности организма изучает наука
– синаптология. Она исследует различные свойства синапсов в организме и их реакции на разнообразные воздействия.
Классификация синапсов. В соответствии с морфологическим принципом синапсы
(рис. 4.9) подразделяются на /6/:
 аксо-аксональные (между двумя аксонами);
аксо-аксональные (между двумя аксонами);
 аксо-дендритические (между аксоном одного нейрона и дендритом другого);
аксо-дендритические (между аксоном одного нейрона и дендритом другого);
 аксо-соматические (между аксоном одного нейрона и телом другого);
аксо-соматические (между аксоном одного нейрона и телом другого);
 дендро-дендритические (между дендритами двух или нескольких нейронов);
дендро-дендритические (между дендритами двух или нескольких нейронов);
 нервно-мышечные (между аксоном нейрона и мышечным волокном);
нервно-мышечные (между аксоном нейрона и мышечным волокном);
 аксо-эпителиальные (между секреторным нервным волокном и грандулоцитом);
аксо-эпителиальные (между секреторным нервным волокном и грандулоцитом);
 межнейронные (общее название синапсов между какими-либо элементами двух нейронов).
межнейронные (общее название синапсов между какими-либо элементами двух нейронов).
Кроме того, все синапсы делят на центральные (нейронейрональные в головном и спинном мозге) и периферические (нервно-мышечные, аксоэпителиальные и синапсы
63
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
вегетативных ганглиев).
В соответствии с нейрохимическим принципом синапсы классифицируют по виду химического вещества – медиатора, с помощью которого происходит возбуждение и торможение эффекторной клетки. Различают адренергические, холинергические, серотонинергические, ГАМК-ергические и др. синапсы.
Сложный синапс на шипике
|
|
|
Ответвление |
Дендриты |
||||||
|
|
|
шипика |
|
|
|
|
|
|
|
|
|
|
|
Шипики |
||||||
Шипик |
|
|||||||||
|
|
|
|
Микро- |
||||||
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
трубочки |
|||
|
|
|
|
|
|
|
Мито- |
|||
|
|
|
|
|
|
|
хондрия |
|||
|
|
|
|
|
|
|
|
Синапти- |
||
|
|
|
|
|
|
|
|
ческие |
||
|
|
|
|
|
|
|
|
пузырьки |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Шипик |
||
|
|
|
|
|
|
Дендрит |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
Аксодендритический синапс |
|||||
|
Тело |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ней- |
Аксон |
||||||||
|
рона |
|||||||||
|
|
|
|
Аксон |
|
Аксосоматические синапсы |
|
|
|||
|
|
|
Эффекторная |
||
|
|
|
|
||
|
|
Аксон |
|||
|
|
|
клетка |
||
|
|
|
|
||
Аксо-аксональный синапс
Рис. 4.9. Различные виды синаптических контактов на пирамидной клетке гиппокампа /6/.
По способу передачи информации синапсы подразделяются на три группы. Первую группу составляют синапсы с химической природой передачи посредством медиаторов, например, нервно-мышечные.
Вторую – синапсы с передачей электрического сигнала непосредственно с прена постсинаптическую мембрану – эфапсы, например, синапсы в сетчатке глаза. По сравнению с химическими они отличаются большой скоростью передачи сигнала, высокой надежностью и возможностью двусторонней передачи возбуждения.
Третья группа представлена «смешанными» синапсами, сочетающими элементы как химической, так и электрической передачи.
По конечному физиологическому эффекту, а также по изменению потенциала постсинаптической мембраны, различают возбуждающие и тормозные синапсы. В возбуждающих синапсах в результате деполяризации постсинаптической мембраны
64

ФИЗИОЛОГИЯ НЕРВНЫХ ВОЛОКОН. СИНАПСЫ |
4 |
|
|
генерируется возбуждающий постсинаптический потенциал (ВПСП). В тормозных синапсах в результате гиперполяризации постсинаптической мембраны возникает тормозной постсинаптический потенциал (ТПСП) /6/.
Этапыимеханизмысинаптическойпередачи
Передача возбуждения в химическом синапсе включает в себя синтез и секрецию медиатора, взаимодействие медиатора с рецепторами постсинаптической мембраны, инактивирование медиатора. В целом синапс осуществляет последовательную трансформацию электрического сигнала, поступающего по нервному волокну, в энергию химических превращений на уровне синаптической щели и постсинаптической мембраны, которая затем снова трансформируется в энергию распространяющегося возбуждения в эффекторной клетке /6/.
Синтез медиатора. Некоторые медиаторы (например, ацетилхолин) синтезируются в цитоплазме синаптических окончаний из нормальных компонентов внутри- и внеклеточных жидкостей, поступающих в них путем пиноцитоза. Другие образуются в теле нейрона, в частности в аппарате Гольджи (пластинчатый комплекс), в перикарионе, а затем путем прямого аксонного транспорта поступают в синаптическое окончание и депонируются в везикулах /6/.
По принципу Дейла каждая нервная клетка синтезирует только один медиатор. В синапсах периферической нервной системы имеется два медиатора (ацетилхолин и норадреналин). Ацетилхолин выделяется в нервно-мышечных синапсах соматической нервной системы, ганглионарных синапсах вегетативной нервной системы, нейроорганных синапсах парасимпатической нервной системы и нейроорганных синапсах симпатической нервной системы в потовых железах и кровеносных сосудах скелетных мышц. Норадреналин выделяется в основной части нейроорганных синапсов симпатической нервной системы. В синапсах соматической нервной системы ацетилхолин является возбуждающим медиатором. В нейроорганных синапсах вегетативной нервной системы ацетилхолин и норадреналин могут вызвать и возбуждение и торможение постсинаптических мембран (амбивалентность медиатора), что зависит от свойств постсинаптических рецепторов. Так, один и тот же медиатор (например, ацетилхолин) может активировать как натриевые (в скелетных мышцах), так и калиевые (в сердце) каналы. В первом случае синапсы, выделяющие ацетилхолин, действует как возбуждающие, во втором – как тормозные.
Таким образом, характер изменения проницаемости постсинаптической мембраны (деполяризация и гиперполяризация) зависит не столько от химической природы медиатора, сколько от особенностей строения клеточных рецепторов разных органов. Именно, поэтому один и тот же медиатор (например, ацетилхолин) возбуждает одни органы (скелетные мышцы) и тормозит другие (сердце).
Кроме основного медиатора, пресинаптические окончания выделяют комедиаторы, которые настраивают чувствительность постсинаптических рецепторов к основному медиатору. В адренергических синапсах комедиаторами являются дофамин, АТФ, гонадотропин и пептид Y. В холинергических синапсах комедиатором является вазоинтестинальный полипептид.
В синапсах центральной нервной системы имеется множество медиаторов. Главным
65

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
возбуждающим медиатором является глутамат, с помощью которого в ЦНС передается 80% информации, а также аспартат. Ацетилхолин, норадреналин, дофамин и серотонин настраивают возбудимость нейронов, определяя уровень бодрствования организма и его мотивационно-эмоциональный фон. Ацетилхолин поддерживает тонус нервных центров как при бодрствовании, так и во время сна. Норадреналин дополнительно активирует нервные центры при напряжении организма. Дофамин способствует регуляции движений и мышечного тонуса. Серотонин сдерживает возбуждение нервных центров в необходимых пределах.
Гамма-аминомасляная кислота (ГАМК) и глицин являются тормозными медиаторами.
Ряд биологически активных веществ обладает модулирующим действием на основные медиаторы (нейромодуляторы). Гистамин повышает чувствительность нервных центров к основным медиаторам. Вещество Р разнообразно модулирует действие основных медиаторов и принимает прямое участие в проведении болевого и эмоционального возбуждения. Энкефалины и эндорфины тормозят выделение основных медиаторов при болевом возбуждении.
Секрециямедиатора
Высвобождение медиатора из синаптических пузырьков имеет квантовый характер.
В состоянии покоя оно незначительно, так как в отсутствие деполяризации пресинаптического окончания происходит только случайное столкновение синаптических пузырьков с пресинаптической мембраной. Скорость спонтанной секреции зависит от уровня поляризации пресинаптической мембраны и количества ионов кальция внутри нервного окончания. В результате экзоцитоза в синаптическую щель попадает лишь небольшое количество медиатора. В ответ на кванты медиатора на постсинаптической мембране возникают миниатюрные постсинаптические потенциалы (МПСП), причем беспорядочно, их величина (0,5 мВ) значительно меньше порога возбуждения постсинаптической мембраны, и нужно 200-300 таких МПП, чтобы возникло распространяющееся возбуждение /6/.
При деполяризации пресинаптической мембраны под влиянием нервного импульса высвобождение медиатора резко усиливается.
Взаимосвязь между деполяризацией пресинаптической мембраны и высвобождением медиатора выполняют ионы кальция, которые всякий раз, когда возбуждение (ПД) достигает пресинаптической части, на короткое время, но в значительных количествах входят в цитоплазму пресинаптического окончания и инициируют высвобождение медиатора из везикул.
И если деполяризация связана с активацией натриевых, то секреция – только кальциевых каналов.
Сложные взаимодействия между электрическим импульсом и синаптическими пузырьками (в присутствии ионов кальция, АТФ и мембранной АТФ-азы) приводят к склеиванию везикул с пресинаптической мембраной и опорожнению медиатора в синаптическую щель (экзоцитоз) (рис. 4.10). Скорость экзоцитоза зависит от скорости высвобождения кальция из эндоплазматической сети пресинаптического окончания и его поступления извне и активности метаболических процессов. Она увеличивается при введении лекарственных средств, содержащих кальций, а также АТФ, биогенных
66
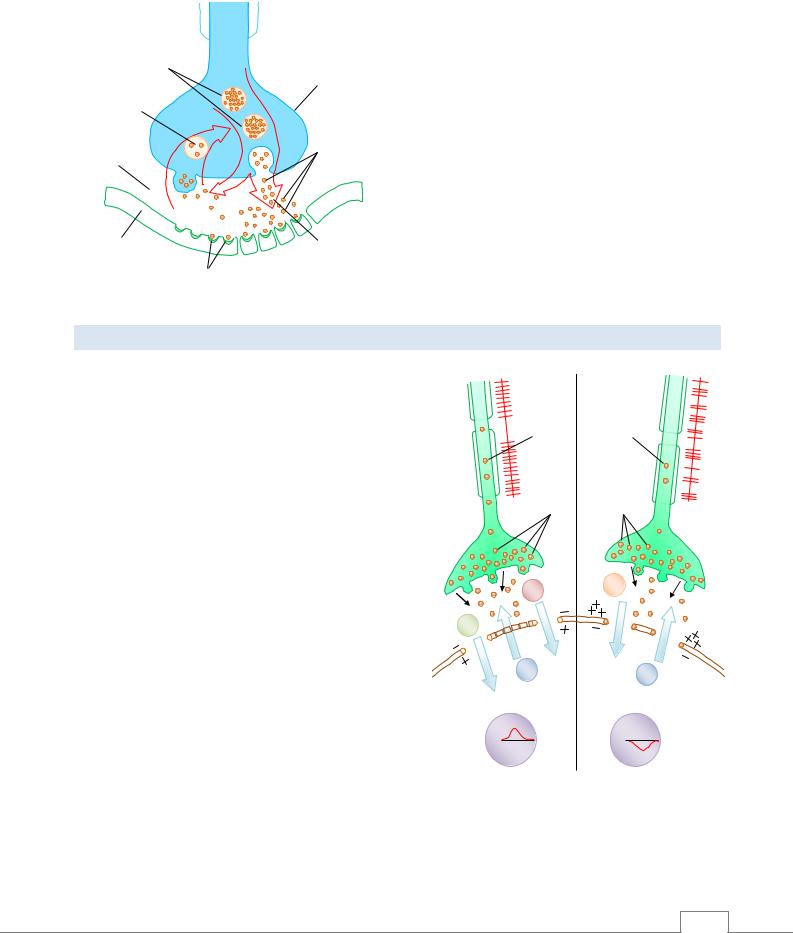
ФИЗИОЛОГИЯ НЕРВНЫХ ВОЛОКОН. СИНАПСЫ |
4 |
|
|
стимуляторов (алоэ, женьшень, боярышник), гормонов /6/.
Синаптические |
Пресинаптическая |
|
|
пузырьки |
мембрана |
|
|
Пиноцитозный |
|
|
|
пузырек |
|
Рис. 4.10. Экзо- и пиноцитоз медиатора в синапсе |
|
Синаптическая |
Молекулы |
||
(схема) /6/. |
|||
щель |
медиатора |
||
|
|||
Постсинаптическая |
Экзоцитоз |
|
|
медиатора |
|
||
мембрана |
|
||
|
|
||
Белки-хеморецепторы |
|
|
Взаимодействиемедиаторасрецепторамипостсинаптическоймембраны
Структурными элементами мембранного рецептора являются два белковых функциональных центра связывания молекул медиатора и пронизывающий мембрану ионоселективный канал. Связь между рецептором и ионоселективным каналом заключается в том, что при изменении (конформации) рецептора осуществляется «запуск» работы ионоселективного канала.
Вследствие конформационных процессов в области белковых ионоселективных каналов изменяется их проницаемость для различных ионов
(рис. 4.11).
При увеличении проницаемости натриевых каналов усиливается поступление этих ионов в клетку с последующей деполяризацией химически возбудимых участков мембраны и возникновением ВПСП, величина которых пропорциональна количеству выделяющегося медиатора. Из множества ВПСП вследствие их суммации возникает распространяющееся возбуждение в соседнем электровозбудимом участке мембраны (потенциал действия). Такие синапсы называются
возбуждающимися (рис. 4.12-I).
 Пресинаптические
Пресинаптические  окончания
окончания
Везикулы с медиатором
Ca2+ Cl-
Na+
K+ |
+ |
|
K |
ВПСП ТПСП
мВ |
мВ |
Возбуждение Торможение
Рис. 4.11. Возбуждающий и тормозной синапсы /1, 4, 6 /.
Если повышается проницаемость калиевых каналов и каналов для хлора, наблюдается
67
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
избыточный выход калия из клетки с одновременной диффузией в нее хлора, что приводит к гиперполяризации мембраны, снижению ее возбудимости и развитию ТПСП. Передача нервных импульсов затрудняется или совсем прекращается. Такие синапсы называются тормозными (рис. 4.12-II).
|
I |
|
II |
|
Аксоплазматический |
Проведение |
Аксоплазматический |
Проведение |
|
ток медиатора |
ток медиатора |
|||
|
|
возбуждения |
|
возбуждения |
Медиатор |
|
|
Медиатор |
|
Везикулы |
|
Na+ |
Везикулы |
Na+ |
|
K+ |
Ca2+ |
K+ |
Ca2+ |
А |
|
|
|
|
Пресинаптическая |
|
ПресинаптическаяCa2+Ca2+ Ca2+ |
||
мембрана |
Ca2+Ca2+ Ca2+ Синаптическая |
|||
|
|
щель |
мембрана |
Синаптическая |
|
|
|
||
|
|
|
щель |
|
|
|
|
|
|
Постсинаптическая |
|
Постсинаптическая |
|
|
мембрана |
|
|
|
|
|
|
мембрана |
|
|
Б |
|
|
|
|
|
|
|
|
|
|
Na+ Na+ Na+ |
Cl– Cl– Cl– |
||
|
|
ВПСП |
|
ТПСП |
|
|
|
|
|
|
|
|
|
Торможение |
В |
|
|
|
возбуждения |
|
|
|
|
|
Потеря части |
|
Пресинаптические |
|
|
|
рецепторы |
|
|
|
медиатора |
|
|
|
|
|
|
|
|
Отсутствие |
|
|
Проведение |
|
проведения |
|
|
|
возбуждения |
|
Г |
|
возбуждения |
|
|
|
|
|
||
|
|
|
|
|
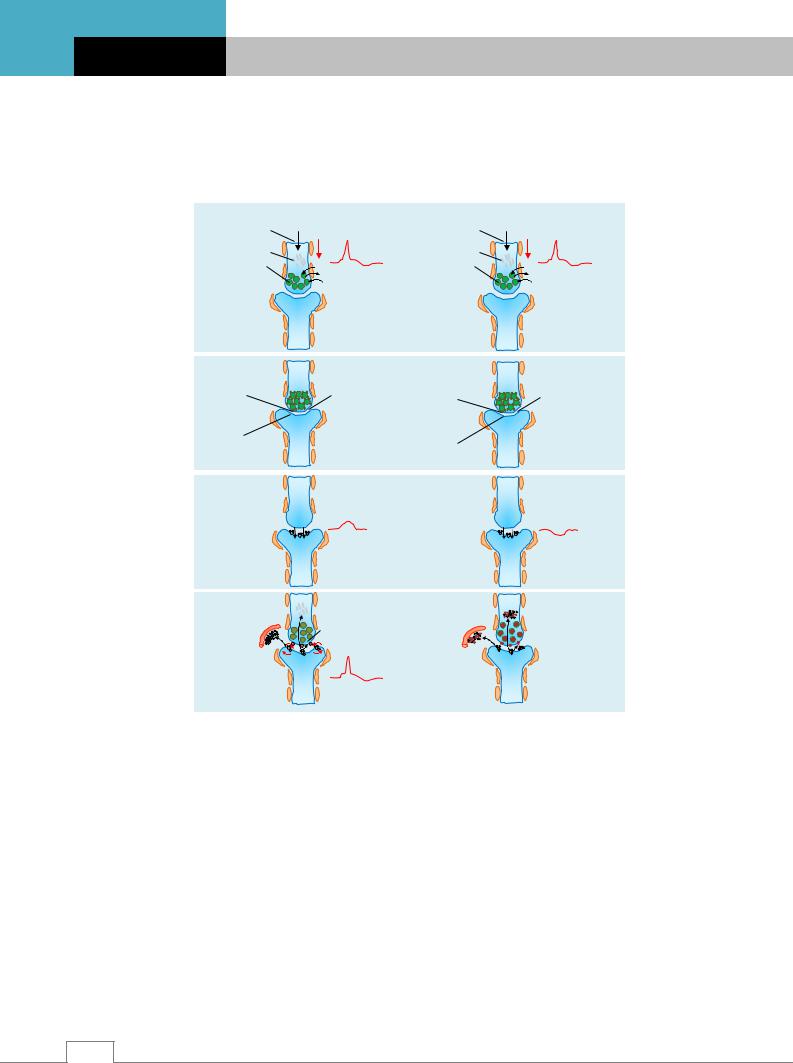
Рис. 4.12. Проведение возбуждения через возбуждающий синапс (I) и торможение проведения возбуждения в тормозном синапсе (II): А – проведение возбуждения по аксону и увеличение проницаемости мембраны для ионов Са2+; Б – Са2+ вызывает движение везикул к пресинаптической мембране. Медиатор выходит в синаптическую щель; В – медиатор присоединяется к рецепторам постсинаптической мембраны и возникает ВПСП или ТПСП; Г – локальные токи способствуют возникновению ПД в соседних с постсинаптической мембраной участках, и возбуждение проводится в возбуждающем синапсе. В тормозном синапсе возбуждение не проводится. Медиатор расщепляется ферментативно, и большая часть продуктов его расщепления активно всасывается обратно в пресинаптическое окончание. Пресинаптические рецепторы оценивают количество выделившегося медиатора /5/.
Таким образом, взаимодействие медиатора с рецепторами может вызывать возбуждение или торможение эффекторной (постсинаптической) клетки, в частности, нейрона; сокращение или расслабление мышечной клетки; образование и выделение гормонов и других биологически активных веществ секреторными клетками. Рецептор «переводит»
68

ФИЗИОЛОГИЯ НЕРВНЫХ ВОЛОКОН. СИНАПСЫ |
4 |
|
|
сообщение, закодированное в молекулярной структуре медиатора, в специфическую физиологическую реакцию.
В возбуждении постсинаптической мембраны, помимо медиаторов, участвуют и другие выделяющиеся в пресинаптической области вещества: ионы кальция, отдельные олигопептиды, такие как вещество Р, факторы роста нервов, оксид азота и др.
Классификация рецепторов постсинаптической мембраны. Рецепторы,
взаимодействующие с ацетилхолином, называются холинорецепторами. В функциональном отношении они подразделяются на две группы – мускарино- (М) и никотино- (Н) чувствительные. В синапсах скелетных мышц и вегетативных ганглиях присутствуют только Н-холинорецепторы, тогда как в сердце и гладких мышцах внутренних органов – преимущественно М-холинорецепторы.
Рецепторы, взаимодействующие с норадреналином, называются адренорецепторами. В функциональном отношении они делятся на α- и β-адренорецепторы. В постсинаптической мембране гладких мышечных клеток внутренних органов и кровеносных сосудов часто соседствуют оба вида адренорецепторов. Действие норадреналина в ряде случаев является деполяризующим, если он взаимодействует с α-адренорецептором, или тормозным – при взаимодействии с β-адренорецепторами. Результатом взаимодействия норадреналина с α-адренорецептором является, например, сокращение мышечной оболочки стенок кровеносных сосудов (артериол), а с β2-адренорецепторами – их расслабление.
Холино- и адренореактивные структуры находятся во всех внутренних органах, железах внутренней и внешней секреции, скелетных и гладких мышцах, вегетативных ганглиях, ЦНС.
Рольхолинэстеразывпроведениивозбуждения
В области нервно-мышечного соединения в больших концентрациях присутствует фермент холинэстераза, способная быстро расщеплять ацетилхолин, выделяющийся в нервном окончании. В естественных условиях к мышце поступают быстро следующие друг за другом нервные импульсы и постсинаптическая мембрана, деполяризованная предшествующей порцией ацетилхолина, становится малочувствительной к действию следующей порции. Чтобы идущие друг за другом нервные импульсы могли осуществлять нормальное возбуждающее действие, необходимо к моменту прихода каждого из них «убрать» предшествующую порцию медиатора. Эту функцию выполняет холинэстераза. Холин, освобождающийся при расщеплении молекул ацетилхолина, переносится обратно в нервное окончание специальной транспортной системой, существующей в пресинаптической мембране.
Фармакологические вещества, обладающие способностью угнетать активность холинэстеразы, называются ингибиторами. К ним относятся: эзерин, простигмин, галантамин. Если на нервно-мышечное соединение действует какое-либо из этих веществ, постсинаптический потенциал увеличивается по своей амплитуде и резко растягивается во времени.
При действии ингибитора холинэстеразы ритмическое раздражение нерва вызывает выраженную суммацию потенциалов концевой пластинки (ПКП), что ведет к стойкой
69

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
деполяризации постсинаптической мембраны и блоку проведения импульсов с нервного волокна на мышечное. При этом стойкая деполяризация постсинапической мембраны приводит соседние участки мышечного волокна в состояние угнетения, обусловленное инактивацией натриевой, и стойким повышением калиевой проницаемости мембраны (состояние «катодической депрессии»).
В отсутствие ингибиторов холинэстеразы при условии частого раздражения нерва постсинаптические потенциалы, вызываемые каждым нервным импульсом, суммируются, поскольку в межимпульсный интервал холинэстераза не успевает полностью расщепить выделяющийся в нервном окончании ацетилхолин. В результате суммации потенциалов постсинаптическая мембрана все более и более деполяризуется.
Химическаяиэлектрическаятеориивозбуждениясинапсов
Согласно химической теории передача возбуждения с нервного волокна на скелетное мышечное волокно представляется следующим образом. При поступлении к пресинаптической области распространяющегося процесса возбуждения в пресинаптической мембране открываются потенциалзависимые кальциевые каналы. Под влиянием Са2+ пресинаптические везикулы с ацетилхолином начинают массивно концентрироваться около пресинаптической мембраны. Как только везикулы соприкасаются с пресинаптической мембраной, происходит лизис везикулярной и пресинаптической мембран – и содержимое везикулы изливается путем экзоцитоза в синаптическую щель. Затем медиатор ацетилхолин взаимодействует с Н-холинорецептором натриевого ионного канала в результате открываются активационные (m) ворота натриевых ионных каналов (рис. 4.13).
АЦХ Рецептор
АЦХ

 Ворота
Ворота
А |
Б |
Рис. 4.13. Схематическое изображение хемовозбудимого ионного канала, активизируемого ацетилхолином. Канал образован макромолекулой белка, погруженной в липидной бислой мембраны. Ворота канала управляются хеморецептором (заштрихован). До взаимодействия молекулы ацетилхолина (АЦХ) c рецептором ворота закрыты (А). Они открываются при связывании АЦХ с рецептором (Б).
При этом ионы Na+ устремляются внутрь постсинаптической клетки, что приводит к деполяризации постсинаптической мембраны и генерации потенциала действия. Схематически процесс передачи возбуждения с нервного волокна на скелетное мышечное волокно можно представить в виде следующей цепи явлений:
нервный импульс → поступление Са2+ внутрь нервного окончания → освобождение из нервного окончания ацетилхолина → взаимодействие ацетилхолина с холинорецептором → активация хемовозбудимых каналов постсинаптической мембраны в результате взаимодействия медиатора с холинорецептором → возникновение ПКП → критическая деполяризация околосинаптической электровозбудимой мембраны → генерация ПД → ПД распространяется по мембране мышечного волокна, вызывая его сокращение.
70