

Нормальная физиология (Пособие для резидентуры)
.pdfНЕЙРОН. КЛАССИФИКАЦИЯ НЕЙРОНОВ |
5 |
|
|
проприоцептивную чувствительность жевательных мышц /3/.
Псевдоуниполярные нейроны имеют один короткий отросток, который разделяется на некотором расстоянии от сомы на два длинных – дендрит и аксон. К псевдоуниполярным относятся нейроны сенсорных ганглиев спинного мозга. Они обеспечивают восприятие болевой, температурной, тактильной чувствительности и др.
Биполярные нейроны имеют один дендрит и один аксон. Этот вид нейронов встречается в периферическом отделе зрительного, обонятельного и слухового анализаторов.
Мультиполярные нейроны имеют один аксон и несколько дендритов, это наиболее распространенный вид нейронов. К ним относятся мотонейроны спинного мозга.
Функции афферентных нейронов тесно связаны с функциями рецепторов. Рецепторы представляют собой специализированные образования, воспринимающие определенные виды раздражений. Рецепторы делят на группы: механо-, термо-, хемо-, фоторецепторы и др. Реакция рецепторного нейрона, предназначенного для передачи информации из области восприятия, имеет 5 стадий:
 преобразование сигнала внешнего раздражения;
преобразование сигнала внешнего раздражения;
 генерация рецепторного потенциала;
генерация рецепторного потенциала;
 распространение рецепторного потенциала по нейрону;
распространение рецепторного потенциала по нейрону;
 возникновение генераторного потенциала;
возникновение генераторного потенциала;
 генерация нервного импульса.
генерация нервного импульса.
Рецепторные нейроны, медленно адаптирующиеся к раздражению, т.е. длительное время генерирующие ПД, называются тоническими; рецепторы, быстро адаптирующиеся к раздражению и короткореагирующие на стимуляцию группой импульсов, называются фазиче-
скими.
Процессы возбуждения нейронов объясняют электрическая и химическая теории.
Электрическая теория возбуждения нейронов. Большинство синапсов расположено на дендритах нейронов и только незначительная часть – на телах нейронов. Синапсы на дендритах получили название интеграторных, а на теле клетки ближе к выходу аксона – детонаторных. Считалось, что мембрана дендритов электрически невозбудима. В настоящее время доказано, что отдельные ее участки обладают потенциалоуправляемой проводимостью, обусловленной наличием кальциевых каналов, активация которых приводит к кальциевым потенциалам действия /1, 4/. У места отхождения, аксонов располагается так называемый «светлый аксонный холмик», или «светлое пятнышко», где синапсы отсутствуют. Этот участок нейрона положительно заряжен по отношению к другим участкам мембраны нейронов. При возбуждении синапсов, располагающихся на дендритах и теле нейронов, увеличивается разность потенциалов между «аксонным холмиком» и остальной частью нейрона. При достижении определенной величины это порождает спайковую активность аксона. Чем круче нарастает разность потенциалов тела нейрона и «аксонного холмика», тем чаще выходная импульсация аксона. Все это порождает определенную конфигурацию импульсации аксона
(рис. 5.3).
81

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
ТПСП
ВПСП
ВПСП
Частота импульсации
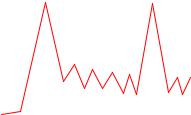

Рис. 5.3. Разность потенциалов между телом нейрона и «аксонным холмиком» порождает выходную импульсацию нейрона /1, 4/.
Химическая теория возбуждения нейронов. Химическая теория связывает процессы возбуждения нейронов со спецификой выделяемых синапсами нейронов медиаторов, олигопептидов и других биологически активных веществ.
Большинство медиаторов могут оказывать двоякое действие – возбуждающее и тормозящее. Только возбуждающее действие оказывают глутамат, аспартат и др., только тормозящее
– ГАМК, глицин.
Возбуждающие медиаторы деполяризуют постсинаптическую мембрану и формируют ВПСП.
Тормозные медиаторы гиперполяризуют постсинаптическую мембрану и формируют на постсинаптической мембране ТПСП.
Медиаторные функции олигопептидов и аминокислот. Олигопептиды – опиоиды, ве-
щество П, ангиотензин II, вазопрессин, окситоцин, а также цитокины модулируют действие медиаторов в синапсах ЦНС. Самостоятельную медиаторную функцию выполняют аминокислоты – глутаминовая кислота, аспарагиновая кислота, ГАМК и глицин. У ряда нейронов ЦНС выявлены специфические рецепторы к глутамату (NMDA-рецепторы, аффинные к N-метил-D-аспартату).
ГАМК – самый распространенный тормозной медиатор в ЦНС. Другой тормозной медиатор, точкой приложения которого являются нейроны продолговатого и спинного мозга – глицин.
5.1. Функциинейронов
 Рецепторная. Способность к восприятию информации обеспечивается рецепторами постсинаптических мембран.
Рецепторная. Способность к восприятию информации обеспечивается рецепторами постсинаптических мембран.
 Память нейрона позволяет хранить информацию как генетического так и приобретенного характера.
Память нейрона позволяет хранить информацию как генетического так и приобретенного характера.
 Интегративная. Основная деятельность нейрона заключается во внутриклеточной обработке информации.
Интегративная. Основная деятельность нейрона заключается во внутриклеточной обработке информации.
 Синтезирующая. В теле нейрона идут процессы синтеза биологически активных веществ – нейромедиаторов и нейромодуляторов, первые из которых обеспечивают пере-
Синтезирующая. В теле нейрона идут процессы синтеза биологически активных веществ – нейромедиаторов и нейромодуляторов, первые из которых обеспечивают пере-
82

НЕЙРОН. КЛАССИФИКАЦИЯ НЕЙРОНОВ |
5 |
|
|
дачу сигнала на клетку, а вторые могут влиять на эффективность передачи, во многом определяя ее характер.
 Транспортная. Синтезируемые вещества транспортируются внутри аксона нервной клетки к синаптическим окончаниям. Транспорт осуществляется при помощи специальных фибриллярных белков.
Транспортная. Синтезируемые вещества транспортируются внутри аксона нервной клетки к синаптическим окончаниям. Транспорт осуществляется при помощи специальных фибриллярных белков.
 Генераторная. Внешняя реакция нейрона заключается, в генерации потенциалов действия, которая происходит в начальном сегменте нейрона.
Генераторная. Внешняя реакция нейрона заключается, в генерации потенциалов действия, которая происходит в начальном сегменте нейрона.
 Кодирующая. Нейрон обладает свойством кодирования, т.е. передачи информации в условной форме кода.
Кодирующая. Нейрон обладает свойством кодирования, т.е. передачи информации в условной форме кода.
 Проводниковая. Нейрон обладает свойством проводника, причем потенциалы действия из области начального сегмента распространяются по аксональной мембране к синапсам.
Проводниковая. Нейрон обладает свойством проводника, причем потенциалы действия из области начального сегмента распространяются по аксональной мембране к синапсам.
 Секреторная. Проявляется в выделении медиаторов и некоторых пептидов в окончании аксона.
Секреторная. Проявляется в выделении медиаторов и некоторых пептидов в окончании аксона.
 Трофическая. На рост аксонов и функционирование синаптических связей влияют так называемые трофические факторы и, в частности, фактор роста нервов, который синтезируется клетками-мишенями, захватывается нейронами и ретроградно транспортируется в сому, где влияет на ферментативные процессы, синтез медиаторов и рост аксона
Трофическая. На рост аксонов и функционирование синаптических связей влияют так называемые трофические факторы и, в частности, фактор роста нервов, который синтезируется клетками-мишенями, захватывается нейронами и ретроградно транспортируется в сому, где влияет на ферментативные процессы, синтез медиаторов и рост аксона
/4/.
5.2. МетодыисследованияфункцийЦНС
Методы исследования функций ЦНС делятся на две группы /6/:
1)непосредственное изучение;
2)опосредованное (косвенное) изучение.
Методы непосредственного изучения функций ЦНС подразделяют на:
 морфологические;
морфологические;
 функциональные.
функциональные.
Морфологические методы. К морфологическим методам относятся макроанатомические и микроскопические исследования строения мозга. На основании этих методов составлены цитоархитектонические карты различных отделов мозга и схемы проводящих путей в ЦНС.
К морфологическим методам относят и метод меченых атомов. Сущность его заключается в том, что вводимые в организм радиоактивные вещества интенсивнее проникают в те нервные клетки мозга, которые в данных момент наиболее функционально активны.
Функциональные методы. К ним относятся:
 разрушение структур ЦНС;
разрушение структур ЦНС;
 раздражение структур ЦНС;
раздражение структур ЦНС;
 стереотаксический метод;
стереотаксический метод;  электрофизиологические методы.
электрофизиологические методы.
83

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Метод разрушения. При сопоставлении разрушения определенных структур мозга с нарушениями конкретных физиологических функций выявлены достоверные связи.
Так, повреждение в области продолговатого мозга приводит к нарушению дыхания, глотания, деятельности сердца и изменению тонуса сосудов. Клиническая практика дает физиологу возможность сопоставить нарушения функций организма с повреждением различных образований мозга вследствие травм, кровоизлияний, опухолей и др. В клинике для диагностики повреждений мозга различного происхождения (опухоли, инсульт и др.) у человека используют методы компьютерной рентгенотомографии, позитронно-эмиссионной томографии, эхоэнцефалографии, ядерного магнитного резонанса и др.
Метод раздражения структур мозга позволяет установить пути распространения возбуждения от места раздражения к органу или ткани, функция которых при этом изменяется. В качестве раздражающего фактора чаще всего применяют электрический ток, который легко дозируется и не оставляет необратимых изменений. В эксперименте на животных применяют метод самораздражения различных участков мозга.
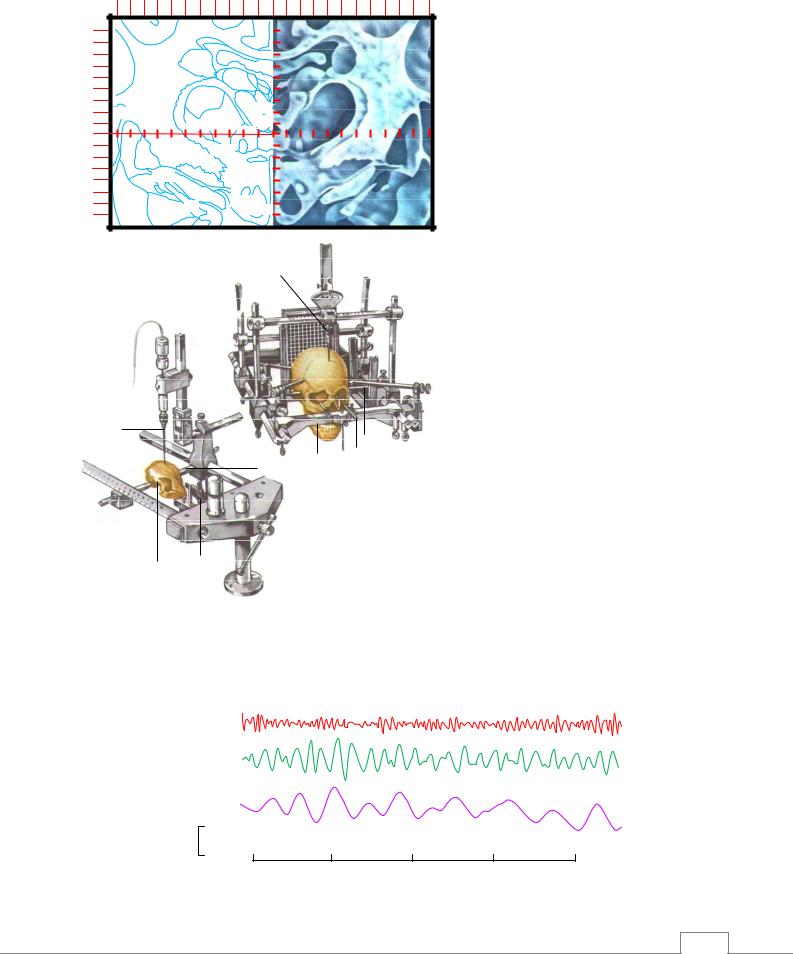
Стереотаксический метод. Высокая точность введения электрода в конкретные образования мозга возможна благодаря стереотаксическим атласам (рис. 5.4). Для фиксации черепа в системе этих плоскостей используют стереотаксический аппарат (рис. 5.4Б), конструкция которого строго соответствует черепу конкретного животного или человека.
Электрофизиологические методы исследования ЦНС включают методы регистрации колебаний сопротивления проходящему через ткань электрическому току (реоэнцефалография), а также суммарной электрической активности мозга (электроэнцефалография); вызванных потенциалов мозга; электрических процессов в отдельных нервных клетках (микроэлектродный метод) /6/.
Электроэнцефалография (ЭЭГ). При экспериментах на животных регистрируется ЭЭГ коры и подкорковых структур. В клинических исследованиях ЭЭГ отводится с помощью электродов, расположенных на интактных покровах головы человека и в некоторых экстракраниальных точках. Отведение ЭЭГ может быть монополярным, при котором активный электрод располагается над определенным участком мозга, а пассивный (референтный) – на мочке уха, подбородке или на носу. При биполярном отведении регистрируется изменение разности потенциалов между электродами, расположенными над мозгом. Основными характеристиками ЭЭГ являются частота и амплитуда. Частота определяется количеством колебаний в секунду и выражается в герцах (Гц). Амплитуда измеряется от пика предшествующей волны до пика последующей волны в противоположной фазе и выражается в микровольтах (мкВ). Определенный тип электрической активности, соответствующий тому или иному состоянию мозга, соответствует понятию «ритм» /4/. Для взрослого бодрствующего человека наиболее характерны α-, β-ритмы (рис. 5.5).
Альфа-(α)-ритм имеет частоту 8-13 Гц, амплитуду до 100 мкВ. Ритм лучше всего выражен в затылочных областях, наибольшую амплитуду имеет в состоянии спокойного расслабленного бодрствования, особенно при закрытых глазах.
Бета-(β)-ритм. Частота 14-40 Гц, амплитуда до 15 мкВ. Ритм появляется при повышении функциональной активности мозга, например, при открывании глаз (реакция десинхронизации).
Тета-(θ)-ритм частотой 4-6 Гц и амплитудой от 40 до 300 мкВ.
84
НЕЙРОН. КЛАССИФИКАЦИЯ НЕЙРОНОВ |
5 |
|
|
|
11 10 9 |
8 |
7 |
6 |
5 |
4 |
3 |
2 |
|
1 |
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 10 11 |
9 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
8 |
|
|
|
|
|
|
|
|
|
CC |
|
|
|
|
|
|
|
|
|
|
7 |
|
|
|
Cd |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
6 |
|
|
|
|
|
|
|
Fx |
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
ST |
AV |
|
|
|
|
|
|
|
|
|
|
|
||||
5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
4 |
|
|
|
|
VA |
|
|
AMRTT |
|
|
|
|
|
|
|
|
|
|
||
3 |
CL |
|
|
R |
|
|
|
|
|
|
|
|
|
|
|
|
||||
2 |
|
|
|
|
VL |
|
|
AM |
|
|
|
|
|
|
|
|
|
|
||
1 |
|
|
|
VPL |
|
|
|
|
|
|
|
|
|
|
|
|
||||
0 |
|
|
|
|
|
|
|
VM |
RE |
|
|
|
|
|
|
|
|
|
||
–1 |
Pu |
|
Cl |
|
LME |
|
|
|
|
|
|
|
|
|
|
|
||||
–2 |
GP |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
–3 |
Acl Acm |
|
En |
|
HL aHa |
|
|
|
|
|
|
|
|
|
|
|||||
–4 |
TO |
|
|
Fx |
|
|
|
|
|
|
|
|
|
|
|
|
||||
–5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
–6 |
Al |
Abm |
ST |
|
MFB |
|
|
|
|
|
|
|
|
|
|
|
|
|||
–7 |
|
|
NHvm |
|
|
|
|
|
|
|
|
|
||||||||
A |
|
|
|
AbpAco |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
1
2
4 3
2
Рис. 5.4. Стереотаксическая техника как метод исследования функций ЦНС.
А – фронтальный срез мозга кошки с осями координат и буквенными обозначениями подкорковых структур мозга;
Б – стереотаксическая установка для проведения опытов на животных и нейрохирургических операций на мозге человека. 1 – электрод; 2 – ушные держатели; 3 – фиксаторы верхней челюсти; 4
– фиксаторы нижней челюсти.
4 3
Б
α-ритм 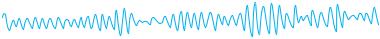
β-ритм
θ-ритм
-ритм |
|
25 мкВ |
1 с |
|
Рис. 5.5. Ритмы электроэнцефалограммы /4/.
85
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Дельта-( )-ритм – медленный, высокоамплитудный, имеющий частоту 0,5-3 Гц и амплиту-
ду 100-150 мкВ характерен для состояния глубокого сна и опухолевых заболеваний головного мозга /4/ .
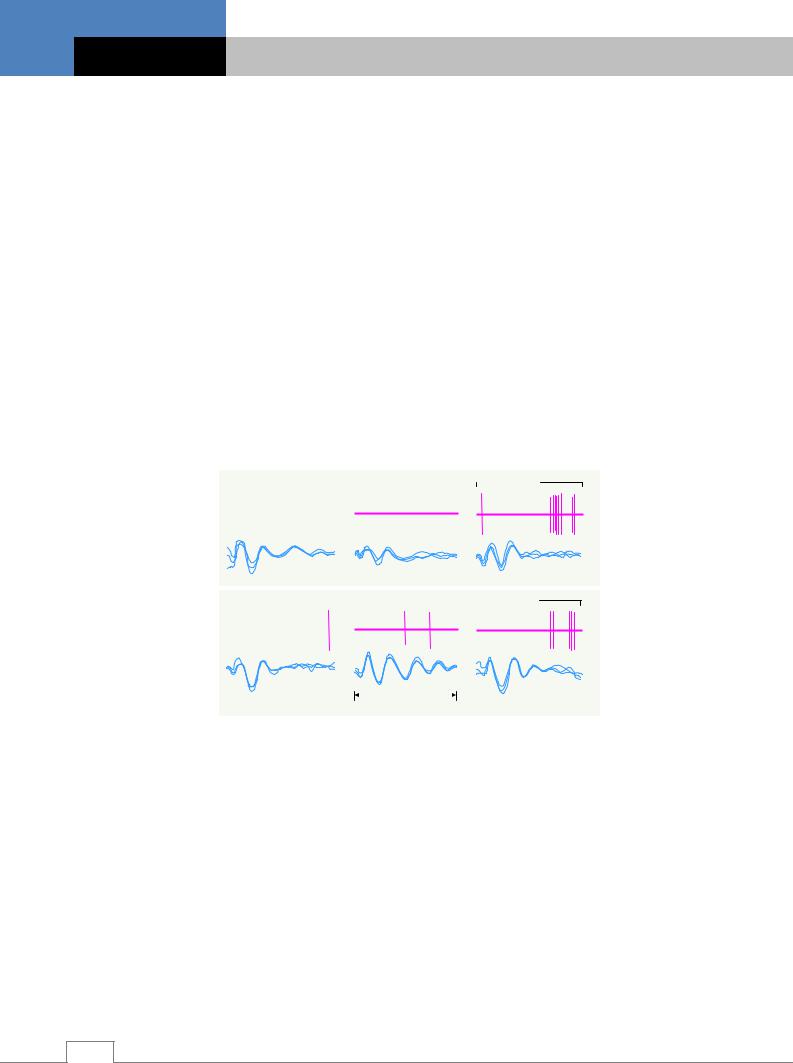
Вызванные потенциалы (ВП). Вызванный потенциал представляет собой колебания биоэлектрического потенциала в структурах головного и спинного мозга при поступлении к ним импульсов возбуждения при раздражении рецепторов, афферентных нервов или других структур.
ВП различаются по конфигурации отдельных компонентов, латентному периоду возникновения, локализации и изменчивости под воздействием различных факторов (рис. 5.6). например, на свет, электрогенное болевое раздражение и их сочетание наибольшую амплитуду и короткий латентный период ВП имеют в фокусе максимальной активности структур головного мозга, куда поступают афферентные импульсы от рецепторов. К таким структурам относятся проекционные зоны коры большого мозга (зрительная, слуховая, соматосенсорная и др.) и специфические ядра таламуса (наружное и внутреннее коленчатые тела и заднее вентральное ядро). В этих областях ВП называются первичными ответами, имеющими амплитуду 400-600 мкВ и латентный период 10-20 мс и состоящими из первично-положительной и первично-отрицательной фаз /6/.
|
|
свет |
|
|
|
|
|
|
|
(свет + ЭКР) |
|
|
|
|
|
|
|
свет |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
А
|
|
|
свет |
|
|
|
|
|
свет |
|
|
|
|
|
|
|
свет |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 с
Б
Рис. 5.6. Вызванные потенциалы и импульсная активность нейрона зрительной области коры большого мозга кролика на световые и электрокожные (болевые) раздражения в процессе формирования условной оборонительной реакции. Хорошо видны первичный (быстрый) ответ и поздние позитивно-негативные колебания вызванных потенциалов на свет. А – подавление поздних ВП при сочетании светового раздражителя с электрокожным – ЭКР (болевым); Б – «облегчение» медленных колебаний ВП в виде их дублирования в ответ на экс-
тренную отмену болевого раздражения /6/.
Фазы первичного ответа имеют преимущественно сенсорное происхождение: положительная фаза отражает постсинаптические процессы на телах нейронов, а отрицательная фаза является выражением местного возбуждения дендритных разветвлений. Таким образом, сенсорные компоненты первичного ответа отражают поступление в головной мозг информации о физических параметрах стимула: интенсивности, временных характеристиках, топографии по отношению к рецептивному полю и др. Возникновение первичного положительного по-
86
НЕЙРОН. КЛАССИФИКАЦИЯ НЕЙРОНОВ |
5 |
|
|
тенциала связывают с возбуждением аксосоматических синапсов на нейронах 3-4 слоя коры, а первичного отрицательного потенциала – с возбуждением аксодендритных синапсов.
Метод вызванных потенциалов используется для определения локализации проекционных зон различных анализаторов в коре мозга.
Микроэлектродный метод. Для микроэлектродных исследований электрической активности отдельных нейронов мозга применяют металлические и стеклянные микроэлектроды, заполненные раствором хлористого натрия или калия. Диаметр кончиков микроэлектродов – 0,5-1 мкм. Для металлических электродов используют нихром, вольфрам, платину, золото, свинец.
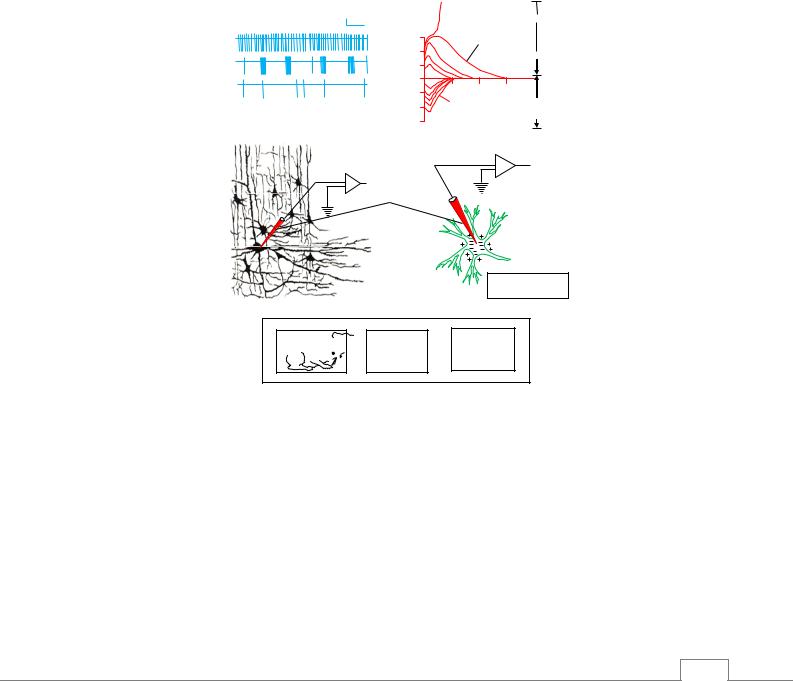
Существуют два способа регистрации клеточной электрической активности: внутриклеточный и внеклеточный (рис. 5.7). При внутриклеточном расположении микроэлектрода регистрируют мембранный потенциал, постсинаптические потенциалы – ВПСП и ТПСП, а также ПД.
1 мВ500 мс |
ПД |
|
Реверсия |
|
|
|
|||
–55 |
ВПСП |
|
||
1 |
|
|
||
2 |
|
|
|
Деполяризация |
|
|
|
|
|
3 |
|
–70 |
1,0 |
1,5 мс |
|
0,5 |
|||
Нейронограмма |
|
ТПСП |
Гиперполя |
|
|
–85 |
|
-ризация |
|
|
|
|
|
|
|
|
Одиночные потенциалы |
||
|
Микроэлектрод |
|
|
|
Внутриклеточное
отведение
Внеклеточное отведение
 Усилитель
Усилитель  Осциллограф
Осциллограф
Рис. 5.7. Микроэлектродный метод исследования нейронной активности. Импульсация на нейронограмме: 1 – смешанная; 2 – пачечная; 3 – одиночная /6/.
Внеклеточный микроэлектрод регистрирует только положительную часть потенциала действия.
Аппликация химических веществ, микроионофорез. Аппликация блокаторов или ак-
тиваторов синаптической передачи может осуществляться непосредственно на кору мозга или путем их подведения через хемотроды к различным глубинным структурам мозга. Вещества могут апплицироваться к отдельным нейронам мозга через микроканюли путем электрического отталкивания зарядов – микроионофорез.
Магнитно-резонансная томография. Этот метод позволяет обнаруживать в головном мозге опухолевые образования и зоны нарушения мозгового кровообращения.
87
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Позитронно-эмиссионная томография. Метод позволяет наблюдать в различных отделах мозга очаги возбуждения при умственной деятельности испытуемых.
Моделирование функций мозга. Построены модели нервных сетей, выполняющие отдельные функции мозга. Создан «детектор интеллекта» (Е.А.Умрюхин), позволяющий определять индивидуальные свойства различных стадий системной организации психической деятельности человека.
Эхоэнцефалография. В основе метода лежит способность ультразвуковых колебаний проникать через кости черепа и ткани головного мозга и отражаться от границ анатомических образований, имеющих разную плотность, благодаря чему можно определять наличие или отсутствие смещения срединных структур мозга.
Магнитоэнцефалография (МЭГ) применяется для выявления области мозга связанных с решением определенных задач, анализа активности данных областей и для анализа изменения их функциональной организации при нарушениях. МЭГ обладает гораздо большей скоростью получения информативного сигнала – в течение 1 мс.
Микродиализ позволяет исследовать химический состав межклеточного пространства в живых тканях и его изменения в процессе жизнедеятельности организма. Принцип микродиализа заключается в диффузии химических соединений, находящихся в межклеточном пространстве, через полупроницаемую мембрану в просвет микродиализного зонда, имплантируемого в исследуемую ткань.
5.3. Нервныецентры
Нервным центром называют совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции той или иной функции. Локализацию нервных центров определяют на основании опытов с раздражением, разрушением, экстирпацией (удалением) или перерезкой различных участков головного или спинного мозга.
Свойстванервныхцентров
Нервные центры обладают рядом характерных свойств, определяемых особенностями синаптического проведения нервных импульсов и структурой нейронных цепей, образующих эти центры.
 Одностороннее проведение. В нервном волокне импульсы могут проводиться в обоих направлениях. В центральной же нервной системе возбуждение может распространяться только в одном направлении: от рецепторного нейрона через промежуточные нейроны к эффекторному. Это явление получило название закона одностороннего проведения возбуждения в нервных центрах. Оно определяет направленность движения нервных импульсов, характерную для рефлекторной дуги.
Одностороннее проведение. В нервном волокне импульсы могут проводиться в обоих направлениях. В центральной же нервной системе возбуждение может распространяться только в одном направлении: от рецепторного нейрона через промежуточные нейроны к эффекторному. Это явление получило название закона одностороннего проведения возбуждения в нервных центрах. Оно определяет направленность движения нервных импульсов, характерную для рефлекторной дуги.
 Замедленное проведение. В нервных центрах проведение возбуждения совершается значительно медленнее, чем в нервных волокнах. Этим объясняется относительная длительность времени рефлекса, т.е. времени от начала раздражения рецептора до появления ответной реакции. Это время называют также латентный период рефлекса. В течение его происходят следующие процессы: возбуждение рецепторов (А), проведение возбуждения по цен-
Замедленное проведение. В нервных центрах проведение возбуждения совершается значительно медленнее, чем в нервных волокнах. Этим объясняется относительная длительность времени рефлекса, т.е. времени от начала раздражения рецептора до появления ответной реакции. Это время называют также латентный период рефлекса. В течение его происходят следующие процессы: возбуждение рецепторов (А), проведение возбуждения по цен-
88

НЕЙРОН. КЛАССИФИКАЦИЯ НЕЙРОНОВ |
5 |
|
|
тростремительным нервным волокнам к нервным центрам (Б), передача возбуждения от одних нейронов на другие внутри центральной нервной системы (В), проведение возбуждения по центральным нервным волокнам от центральной нервной системы (Г), передача возбуждения с нерва на рабочий орган и латентный период его деятельности (Д). Таким образом, время рефлекса (Р) представляет суммарное выражение продолжительности всех этих процессов: Р=А+Б+В+Г+Д. То время, в течение которого происходит внутрицентральное проведение возбуждения с афферентного нейрона на эфферентный (В), носит название истинного, или центрального, времени рефлекса. Для определения его необходимо из общего времени рефлекса вычесть время, затраченное на все остальные перечисленные выше процессы, В=Р–(А+Б+Г+Д). У человека наименьшую продолжительность имеет время сухожильных рефлексов, например коленного: оно составляет всего 20-24 мс. Центральное время этих рефлексов равно 3 мс. Время рефлекса зависит от силы раздражения и от состояния центральной нервной системы. При сильном раздражении оно короче, чем при слабом, при утомлении оно удлиняется, при повышении возбудимости нервных центров значительно укорачивается.
Замедленное проведение возбуждения в центральной нервной системе обусловлено особенностями передачи возбуждения через синапсы. Передача возбуждения через синапс складывается из трех основных процессов:
 выделения медиатора нервным окончанием в ответ на пришедший к нему по аксону импульс;
выделения медиатора нервным окончанием в ответ на пришедший к нему по аксону импульс;
 диффузии медиатора через синаптическую щель к постсинаптической мембране;
диффузии медиатора через синаптическую щель к постсинаптической мембране;
 возникновения под влиянием этого медиатора возбуждающего постсинаптического потенциала. Точные измерения позволили установить, что между моментом поступления импульса к нервному окончанию и началом возбуждающего постсинаптического потенциала проходит примерно 0,5 мс, необходимые для выделения нервным окончанием медиатора и его диффузии к постсинаптической мембране. Это время называют синаптической задержкой. Между моментом возникновения постсинаптического потенциала и появлением пикового потенциала проходит еще 1,5-2 мс – это время, необходимое для нарастания возбуждающего постсинаптического потенциала до критической величины. Таким образом в общей сложности, проведение возбуждения через один синапс требует 2-3 мс.
возникновения под влиянием этого медиатора возбуждающего постсинаптического потенциала. Точные измерения позволили установить, что между моментом поступления импульса к нервному окончанию и началом возбуждающего постсинаптического потенциала проходит примерно 0,5 мс, необходимые для выделения нервным окончанием медиатора и его диффузии к постсинаптической мембране. Это время называют синаптической задержкой. Между моментом возникновения постсинаптического потенциала и появлением пикового потенциала проходит еще 1,5-2 мс – это время, необходимое для нарастания возбуждающего постсинаптического потенциала до критической величины. Таким образом в общей сложности, проведение возбуждения через один синапс требует 2-3 мс.
Исходя из этого, можно определить количество синапсов, через которые проходит нервный импульс в ЦНС. Например, поскольку центральное время большинства сухожильных рефлексов, в том числе коленного рефлекса, у человека составляет примерно, 3 мс, то считают, что дуга этих рефлексов включает только один синапс. Такие рефлексы называются
моносинаптическими.
Зависимость рефлекторного ответа от силы и длительности раздражения. При уси-
лении раздражения рецептивного поля увеличивается число возбужденных рецепторов и нервных волокон, по которым импульсы поступают в нервный центр, а следовательно, возрастает и число промежуточных и эффекторных нейронов, вовлекаемых в реакцию. Вместе с тем увеличивается частота нервных импульсов, возникающих в рецепторах и соответственно в каждом из нейронов, что также приводит к усилению рефлекса (усилению сокращения мышц, усилению секреции железы и т.д.). Увеличение продолжительности раздражения даже при постоянстве силы последнего в ряде случаев также ведет к усилению рефлекса за счет вовлечения в реакцию новых нервных элементов.
89

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ
Суммация возбуждений. По И.М.Сеченову различают два вида суммации: последова-
тельную (временную) и пространственную. Временной суммацией называют взаимодействие возбуждений, приходящих в нервный центр с коротким интервалом друг за другом по одним и тем же афферентным нервным волокнам. Если сила каждого из этих стимулов достаточна для вызова рефлекса, то при их ритмическом применении рефлекс усиливается. Если же подобрать силу раздражения таким образом, чтобы каждый одиночный стимул сам по себе рефлекса не вызывал, то при последовательном применении их друг за другом можно на-
блюдать возникновение рефлекторного ответа. |
|
|
Пространственная суммация возбуждений |
|
|
возникает, когда два или несколько раздражений |
|
|
действуют одновременно на разные рецепторы, |
|
|
относящиеся к одному и тому же рецептивному |
|
|
полю. Так, рефлекс почесывания у собаки можно |
|
|
вызывать, прикладывая одновременно подпоро- |
|
|
говые раздражения к двум участкам кожи, наход- |
|
|
ящимся один от другого на расстоянии 10 см. |
|
|
Каждый такой раздражитель, действуя в отдель- |
1 сек |
|
ности, не вызывает рефлекса почесывания, при |
1 |
|
сочетании же их возникает рефлекторная реакция |
2 |
|
(рис. 5.8). |
Рис. 5.8. Суммация раздражений |
|
Пространственная суммация происходит и в |
||
(по Ч.Шеррингтону): 1 – подпороговое |
||
том случае, если подпороговые раздражения с |
раздражение одного центростремительного |
|
интервалом между ними не больше 15 мсек нано- |
пути; 2 – подпороговое раздражение дру- |
|
сятся на два центростремительных нервных во- |
гого центростремительного пути. Приме- |
|
локна одного и того же рецептивного поля реф- |
ненные в отдельности раздражения 1 и 2 не |
|
лекса. |
дают эффекта. Одновременное применение |
|
Механизм суммации возбуждений в нерв- |
их вызывает рефлекс почесывания. |
ных центрах состоит в следующем. Для возникновения потенциала действия в нейроне необходимо, чтобы деполяризация постсинаптиче-
ской мембраны нервной клетки достигла определенного критического уровня. Такая деполяризация постсинаптической мембраны возникает под влиянием возбуждающего медиатора, выделяющегося нервными окончаниями. Однако порция медиатора, выбрасываемого каждым отдельным нервным окончанием в ответ на одиночный импульс, очень мала, вследствие чего возбуждающий постсинаптический потенциал, возникающий в одиночном синапсе, в 8- 10 раз меньше пороговой величины. Поэтому критическая деполяризация мембраны, необходимая для возникновения распространяющегося возбуждения, возможна либо при одновременном возбуждении многих синапсов, расположенных на одной и той же клетке, либо при поступлении к одному и тому же синапсу серии нервных импульсов, следующих друг за другом с короткими интервалом. И в том, и в другом случае происходит складывание постсинаптических потенциалов. Различие состоит лишь в том, что при одновременном возбуждении нескольких рядом расположенных синапсов постсинаптические потенциалы сумми-
90