

62. Синтез триацилглицеридов и фосфолипидов.
В печени есть фермент глицеролкиназа:
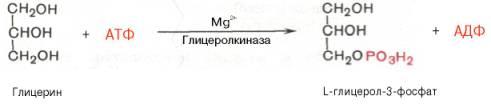
В жировой ткани и
мышцах

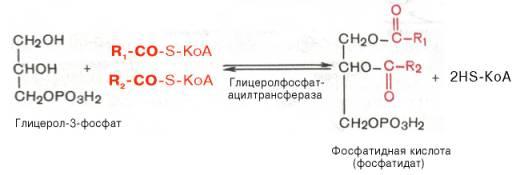
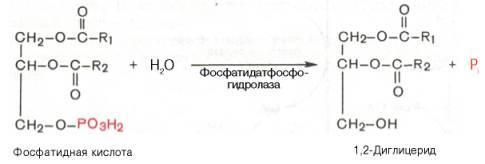
Далее идут присоединения к глицерол-3-фосфату активированных жирных кислот. Образуется фосфатидат.

63. Кетоновые тела и их роль в метаболизме.
Кетоновые тела:
![]() Кетоновые
тела — топливо для мышечной ткани, почек
и др.(особенно при недостатке глюкозы,
голодании) Предотвращают излишнюю
мобилизацию жирных кислот из жировых
депо.[2] Во
время голодания кетоновые тела являются
одним из основных источников энергии
для мозга.[4][5] Печень,
синтезируя кетоновые тела, не способна
использовать их в качестве энергетического
материала (не располагает соответствующими
ферментами).
Синтез и использование:
Кетоновые
тела — топливо для мышечной ткани, почек
и др.(особенно при недостатке глюкозы,
голодании) Предотвращают излишнюю
мобилизацию жирных кислот из жировых
депо.[2] Во
время голодания кетоновые тела являются
одним из основных источников энергии
для мозга.[4][5] Печень,
синтезируя кетоновые тела, не способна
использовать их в качестве энергетического
материала (не располагает соответствующими
ферментами).
Синтез и использование:

64. Физико-химические свойства белков. Изоэлектрическое состояние и изоэлектрическая точка аминокислот и белков.
Наиболее характерными физико-химическими свойствами белков являются высокая вязкость растворов, незначительная диффузия, способность к набуханию в больших пределах, оптическая активность, подвижность в электрическом поле, низкое осмотическое давление и высокое онкотическое давление, способность к поглощению УФ-лучей при 280 нм (это свойство, обусловленное наличием в белках ароматических аминокислот, используется для количественного определения белков).
Белки, как и аминокислоты, амфотерны благодаря наличию свободных NH2- и СООН-групп. Для них характерны все свойства кислот и оснований. В зависимости от реакции среды и соотношения кислых и основных аминокислот белкив растворе несут или отрицательный, или положительный заряд, перемещаясь к аноду или катоду. Это свойство используется при очистке белков методом электрофореза.
Белки обладают явно выраженными гидрофильными свойствами.
Значение рН, при котором белок приобретает суммарный нулевой заряд, называют"изоэлектрическая точка" и обозначают как pI. В изоэлектрической точке количество положительно и отрицательно заряженных групп белка одинаково, т.е. белок находится в изоэлектрическом состоянии.
65.Биохимические механизмы переваривания белков в жкт.
В ротовой полости не происходит.
В желудке главные клетки слизистой оболочки секретируют пепсиноген – предшественник протеолитического фермента пепсина. В результате аутокатализа в кислой среде желудочного сока фермент активируется. Соляная кислота поддерживает рН в пределах 1,5-2,0. Это оптимальные условия для активной работы фермента. В кислой среде белки корма подвергаются денатурации, что делает их более доступными ферментативному протеолизу. Пепсин быстро гидролизует в белках пептидные связи, образованые ароматическими аминокислотами и медленно связи между лейцином и дикарбоновыми аминокислотами.
В тонком отделе кишечника происходит дальнейший гидролиз пептидов до аминокислот. Туда поступает панкреатический сок с рН 7,8-8,2. Он содержит неактивные предшественники протеаз: трипсиноген, химотрипсиноген, прокарбоксипептидазу, проэластазу. Слизистой кишечника вырабатывается фермент энтеропептидаза, который активирует трипсиноген до трипсина, а последний уже все остальные ферменты. Протеолитические ферменты содержатся также в клетках слизистой кишечника, поэтому гидролиз небольших пептидов происходит после их всасывания. Конечный результат действия ферментов желудка и кишечника – расщепление почти всей массы пищевых белков до свободных аминокислот.
Всасывание аминокислот происходит в тонком отделе кишечника. Это активный процесс и требует затраты энергии. Основной механизм транспорта – гамма-глутамильный цикл. В нем участвует 6 ферментов и трипептид глутатион (глутамилцистеинилглицин). Ключевой фермент – гамма-глутамилтрансфераза. Кроме того, процесс всасывания АК требует присутствия ионовNa+. Аминокислоты попадают в портальный кровоток – в печень и в общий кровоток. Печень и почки поглощают аминокислоты интенсивно, мозг избирательно поглощает метионин, гистидин, глицин, аргинин, глутамин, тирозин.
В толстом отделе кишечника не всосавшиеся по каким-либо причинам (недостаток или низкая активность протеолитических фрементов, нарушение процессов транспорта АК) пептиды и АК подвергаются процессам гниения. При этом образуются такие продукты как: фенол, крезол, сероводород, метилмеркаптан, индол, скатол, а также группа соединений под общим названием «трупные яды» - кадаверин, путресцин. Эти вещества всасываются в кровь и поступают в печень, где подвергаются конъюгации с глюкуроновой кислотой и другим процессам обезвреживания (см. подробнее главу "Биохимия печени"). Затем они выводятся из организма с мочой.