

13. Моногибридное скрещивание
Фенотип и генотип. Моногибриднымназывается скрещивание, при котором родительские формы отличаются друг от друга по одной паре контрастных, альтернативных признаков.
Признак —любая особенность организма, т. е. любое отдельное его качество или свойство, по которому можно различить две особи. У растений это форма венчика (например, симметричный—асимметричный) или его окраска (пурпурный—белый), скорость созревания растений (скороспелость—позднеспелость), устойчивость или восприимчивость к заболеванию и т. д.
Совокупность всех признаков организма, начиная с внешних и кончая особенностями строения и функционирования клеток, тканей и органов, называется фенотипом. Этот термин может употребляться и по отношению к одному из альтернативных признаков.
Признаки и свойства организма проявляются под контролем наследственных факторов, т. е. генов. Совокупность всех генов организма называют генотипом.
Примерами моногибридного скрещивания, проведенного Г. Менделем, могут служить скрещивания гороха с такими хорошо заметными альтернативными признаками, как пурпурные и белые цветки, желтая и зеленая окраска незрелых плодов (бобов), гладкая и морщинистая поверхность семян, желтая и зеленая их окраска и др.
Единообразие гибридов первого поколения (первый закон Менделя). При скрещивании гороха с пурпурными и белыми цветками Мендель обнаружил, что у всех гибридных растений первого поколения (F1) цветки оказались пурпурными. При этом белая окраска цветка не проявлялась (рис. 3.1).
Мендель установил также, что все гибриды F1 оказались единообразными (однородными) по каждому из семи исследуемых им признаков.
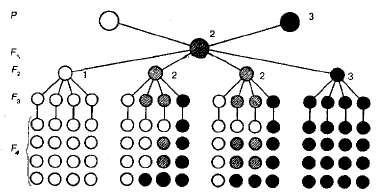
Рис. 3.1. Схема моногибридного скрещивания: I — гомозиготные особи с доминантным признаком; 2 — гетерозиготные особи с доминантным или промежуточным признаком; 3 — гомозиготные особи с рецессивным признаком.
Следовательно, у гибридов первого поколения из пары родительских альтернативных признаков проявляется только один, а признак другого родителя как бы исчезает. Явление преобладания у гибридов F1 признаков одного из родителей Мендель назвал доминированием, а соответствующий признак — доминантным. Признаки, не проявляющиеся у гибридов F1 он назвал рецессивными.
Поскольку все гибриды первого поколения единообразны, это явление было названо К. Корренсом первым законам Менделя, или законом единообразия гибридов первого поколения, а также правилом доминирования.
14. Закон расщепления (второй закон Менделя).Из гибридных семян гороха Мендель вырастил растения, которые подверг самоопылению, и образовавшиеся семена вновь высеял. В результате было получено второе поколение гибридов, или гибриды F2. Среди последних обнаружилось расщепление по каждой паре альтернативных признаков в соотношении примерно 3:1, т. е. три четверти растений имели доминантные признаки (пурпурные цветки, желтые семена, гладкие семена и т. д.) и одна четверть — рецессивные (белые цветки, зеленые семена, морщинистые семена и т. д.). Следовательно, рецессивный признаку гибрида F1 не исчез, а только был подавлен и вновь проявился во втором поколении. Это обобщение позднее было названо вторым законом Менделя, или законом расщепления.
Гомозиготные и гетерозиготные особи. Чтобы выяснить, как будет осуществляться наследование признаков в третьем, четвертом и последующих поколениях, Мендель путем самоопыления вырастил гибриды этих поколений и проанализировал полученное потомство. Он выяснил, что растения, обладающие рецессивными признаками (например, белые цветки), в следующих поколениях (F3 F4 и т. д.), воспроизводят потомство только с белыми цветками (см. рис. 3.1).
Иначе вели себя гибриды второго поколения, обладающие доминантными признаками (например, пурпурными цветками). Среди них при анализе потомства Мендель обнаружил две группы растений, внешне совершенно неразличимых по каждому конкретному признаку.
Первая группа, составляющая 1/3 от общего числа растений с доминантным признаком, далее не расщеплялась, т. е. во всех последующих поколениях у них обнаруживалась только пурпурная окраска цветков. Оставшиеся 2/3 растений второго поколения в F3, снова давали расщепление такое же, как в F2 т. е. на три растения с пурпурными цветками появлялось одно с белыми.
Особи, которые не дают в потомстве расщепления и сохраняют свои признаки в «чистом» виде, называют гомозиготными, а те, у которых в потомстве происходит расщепление, —гетерозиготными.
Таким образом, Менделем впервые было установлено, что растения, сходные по внешним признакам, могут обладать различными наследственными свойствами.
Аллелизм. Для установления причины расщепления, причем в строго определенных численных отношениях доминантных и рецессивных признаков, следует вспомнить, что связь между поколениями при половом размножении осуществляется через половые клетки (гаметы). Очевидно, гаметы несут материальные наследственные задатки, или факторы, определяющие развитие того или иного признака. Эти факторы позже и были названы генами.
В соматических клетках диплоидного организма эти задатки являются парными: один получен от отцовского организма, а другой — от материнского. Мендель предложил обозначать доминантные наследственные задатки заглавной буквой (например,А), а соответствующие им рецессивные задатки прописной буквой (а). Пару генов, определяющих альтернативные признаки, называют аллеломорфной парой, а само явление парности — алле-лизмом.
Каждый ген имеет два состояния — А и а, поэтому они составляют одну пару, а каждого из членов пары называют аллелем. Таким образом, гены, расположенные в одних и тех же локусах (участках) гомологических хромосом и определяющие альтернативное развитие одного и того же признака, называются аллельными. Например, пурпурная и белая окраска цветка гороха является доминантным и рецессивным признаками соответственно двум аллелям (А и а) одного гена. Благодаря наличию двух аллелей возможны два состояния организма: гомо- и гетерозиготные. Если организм содержит одинаковые аллели конкретного гена (АА или аа), то он называется гомозиготным по данному гену (или признаку), а если разные(Аа) — то гетерозиготным. Следовательно, аллель — это форма существования гена.
Примером трехаллельного гена является ген, определяющий у человека систему группы крови АВ0. Аллелей бывает и больше: для гена, контролирующего синтез гемоглобина человека, их известно много десятков.
Статистический анализ расщепления. Представим результаты опытов Менделя по моногибридному скрещиванию гороха в виде схемы (рис. 3.2). Символы Р, F1, F2 и т- д. обозначают родительское, 1-е и 2-е поколение соответственно, знак умножения указывает скрещивание, символ о* обозначает мужской пол, a Q — женский. Из схемы видно, что в родительском поколении (Р) материнская и отцовская формы гомозиготны по исследуемому признаку, поэтому производят гаметы только с аллелем Аили только с а.
При оплодотворении эти гаметы образуют зиготу, которая имеет оба аллеляАа — доминантный и рецессивный. В результате все гибриды F1единообразны по конкретному признаку, поскольку доминантный аллель А подавляет действие рецессивного аллеля а. Во время образования гамет аллели А и а попадают в них по одному. Следовательно, гибридные организмы способны производить гаметы двух типов, несущие аллели А иа, т. е. являются гетерозиготными.
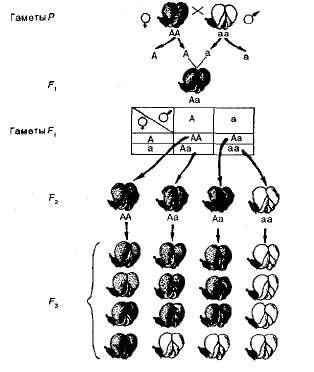
Рис. 3.2. Наследование пурпурной и белой окраски цветков гороха.
Для облегчения расчета сочетаний разных типов гамет английский генетик Р. Пеннет предложил производить запись в виде решетки, которая и вошла в литературу под назван и ем решетка Пеннета (см. рис. 3.2). Слева по вертикали располагаются женские гаметы, сверху по горизонтали — мужские. В квадраты решетки вписывают образующиеся сочетания гамет, которые соответствуют генотипам зигот.
При самоопылении в F2 получается расщепление по генотипу вотношении 1АА:2Аа:1аа, т. е. одна четвертая часть гибридов гомозиготны по доминантным аллелям, половина — гетерозиготны и одна четвертая часть — гомозиготны по рецессивным аллелям. Так как генотипам АА и Аасоответствует один и тот же фенотип — пурпурная окраска цветка, расщепление по фенотипу будет следующим; 3 пурпурных: 1 белый. Следовательно, расщепление по фенотипу не совпадает с расщеплением по генотипу.
Теперь легко объяснить, почему гомозиготные белоцветко-вые растения второго поколения с рецессивными аллелями аа при самоопылении b F3дают только себе подобных. Такие растения производят гаметы одного типа, и, как следствие, расщепления не наблюдается. Ясно также, что среди пурпурноцветковых 1 /3 доминантных гомозигот (АА) также не будет давать расщепления, а 2/3 гетерозиготных растений (Аа) будут давать b F3расщепление 3:1, как и у гибридов F2
На основании, анализа результатов моногибридното скрещивания были сформулированы не только первый и второй законы Менделя и правило доминирования, но и правило чистоты гамет.
15. Влияние одного гена на развитие двух и большего числа признаков называется множественным, или плейотропным, действием, а само явление получило название плейотропии (от греческого pleistos - множественный, наибольший). Биохимическая природа плейотропного действия гена выяснена довольно хорошо. Один белок-фермент, образующийся под контролем одного гена, определяет развитие не только данного признака, но и воздействует на вторичные реакции биосинтеза различных других признаков и свойств, вызывая их изменения. Плейотропия широко распространена: большинство генов у всех организмов обладают множественным действием. Это явление впервые было обнаружено Г. Менделем. Он обнаружил, что у растения с кукурузными цветками одновременно всегда имелись красные пятна в пазухах листьев, а семенная кожура была серого или бурого цвета. Эти три признака определялись действием одного гена. Недавно было установлено, что многим индуцированным мутациям гороха свойственна высокая степень плейотропии, проявляющаяся в изменении до десяти и более признаков. Н. И. Вавилов и О. В. Якушкина, изучая наследование некоторых признаков у персидской пшеницы, выяснили, что доминантный ген черной окраски колоса одновременно вызывает опущение колосковых чешуй. В генотипе человека известны гены, обладающие плейотропным действием. Например, известен ген, вызывающий характерную картину синдрома марфана. Такие люди отличаются длительным ростом конечностей, особенно ног и пальцев рук (паучьи пальцы). К тому же этот ген вызывает дефект в хрусталике глаза. Другим примером плейотропности гена у человека может послужить мутация серповидноклеточности. В этом случае мутация нормального аллеля ведет к изменению в молекулярной структуре белка гемоглобина. В результате мутированные эритроциты теряют способность к транспорту кислорода и вместо нормальной, круглой, приобретают серповидную форму. У людей, гомозиготных по этому признаку, развивается острая анемия, как правило, люди гибнут при рождении. Люди, гетерозиготные по этому аллелю, часто проявляют серповидноклеточность без нарушения транспорта кислорода и при этом обладают повышенной устойчивостью против малярийных комаров. В результате складывается парадоксальная ситуация, в которой ген летален у человека в гомозиготном состоянии, тем не менее он получает широкое распространение. Причиной служит то, что гетерозиготные люди меньше заболевают тропической малярией. В этом случае увеличение идет на гетерозигот, число которых в популяциях больше, чем людей, гомозиготных по этой мутации. Это явление было обнаружено в Средиземноморье и в некоторых других районах. Под взаимодействием генов понимается не непосредственное воздействие одного гена на другой (одного участка молекулы ДНК на другой участок). В действительности взаимодействие генов имеет биохимическую природу. Оно основано на взаимодействии синтезируемых под контролем генов генетических продуктов (РНК, затем белков). Белки могут вступать друг с другом в различные реакции: одни белки подавляют действие других белков или, наоборот, дополняют действие друг друга, могут вызывать различные мутации генов, вследствие чего ген кодирует белок в измененной форме. Все эти взаимодействия между белками, синтезируемым под контролем генов, приводят к формированию организмов, обладающих определенным набором признаков. Известны два вида взаимодействия генов: аллельное и неаллельное. Аллельным называют взаимодействие между аллельными генами, неаллельным - между неаллельными, т. е. между генами, локализованными в разных аллелях.
1. АЛЛЕЛЬНОЕ ВЗАИМОДЕЙСТВИЕ Различают три вида неаллельного взаимодействия: доминирование, неполное доминирование и кодоминирование. 1.1. Доминирование - простейшая форма взаимодействия - была открыта еще Менделем. При доминировании доминантный ген полностью подавляет проявление рецессивного гена. Например, при скрещивании растений гороха с круглыми АА и морщинистыми аа образуются только круглые семена в первом поколении, во втором происходит расщепление в соотношении 3 круглых на 1 морщинистый. Возникает вопрос - почему один ген доминантный, а другой рецессивный? Рецессивный аллель гена может проявиться в результате мутации. Измененный ген либо не кодирует белок, либо кодирует белок меньшей активности. В данном примере рецессивный генотип не влияет на жизнеспособность, но если белок необходим для жизни данного организма, то мутантный ген является летальным. Доминантные летальные аллели существуют в генофонде популяций многих видов. Рецессивные летальные гены не проявляются у гетерозиготного организма. Подсчитано, что человек гетерозиготен по 30 летальным рецессивным генам. Это объясняет тот факт, почему между потомками от браков среди близких родственников наблюдают высокую частоту летальных исходов. 1.2. НЕПОЛНОЕ ДОМИНИРОВАНИЕ При этом типе взаимодействия генов доминирование носит промежуточный характер. Например, при скрещивании красноцветковых растений: А белоцветковых и а львиного зева в первом поколении у гибридов Аа розовая окраска. Во втором происходит расщепление; 1 красноцветковые к 2 розовоцветковым и к 1 белоцветковому. Неполное доминирование объясняется тем, что один ген из аллельной пары не обеспечивает достаточного для нормального проявления его признака белкового продукта. При этой форме взаимодействия генов все гомозиготы и гетерозиготы сильно отличаются друг от друга по фенотипу.
1.3. КОДОМИНИРОВАНИЕ Кодоминированием называют проявление в гетерозиготном состоянии признаков, обусловленных действием обоих аллелей, т. е. каждый из аллельных генов кодирует определенный белок, а в гетерозиготном состоянии проявляются оба. По типу кодоминирования у человека наследуются группы крови. Так, аллели Ia и Ibкодоминанты. У гетерозигот Ia Ib присутствуют оба генных продукта. Аллели Ia Ib доминируют над рецессивной аллелью I. Поэтому у человека система АВО определяет 4 группы крови: A- Ia Ib или IaI0, B-IbIb или IbI0, AB- Ia Ib но группа 0 - i0i0.
16. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ Взаимодействие неаллельных генов проявляется в четырех основных формах: комплементарности, эпистазе, полимерии и модифицирующем действии генов. 2.1. КОМПЛЕМЕНТАРНОЕ ДЕЙСТВИЕ ГЕНОВ Комплементарное (дополнительное) действие наблюдается в случаях, когда неаллельные гены раздельно не проявляют своего действия, но при одновременном присутствии в генотипе обусловливают развитие нового фенотипического признака. В некоторых случаях при соединении дополнительных генов в генотипе возникают признаки, свойственные диким формам. Комплементарное действие наиболее четко проявляется, когда скрещиваются две белые формы некоторых животных (кур) или растений (душистого горошка, белого клевера), а в потомстве появляются окрашенные формы. Разберем комплементарное действие генов на примере душистого горошка. В одном из опытов В. Бэтсона при скрещивании двух форм душистого горошка, имевших белые цветки в первом поколении F1, все гибридные растения оказались с красными цветками. При самоопылении этих растений или скрещивании их между собой в F2 получается расщепление в соотношении: 9 красноцветковых к 7 белоцветковым растениям. Во втором поколении образуются четыре типа гамет, которые при оплодотворении дают 16 типов зигот. В 9 случаях в зиготу попадают оба доминантных гена (А и В), и получаются растения с красными цветками; в шести случаях образуются гаметы, в которые попадает только один доминантный ген из двух взаимодействующих неаллельных пар (А или В); в одном сочетании гамет нет ни одного доминантного гена, и поэтому они дают растения с белыми цветками. Упрощенно механизм этого взаимодействия можно объяснить следующим образом. Предположим, ген А обусловливает синтез красного пропигмента, а ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Поэтому в генотипах, в которых присутствует только один доминантный из взаимодействующей неаллельной пары, пигмента не образуется, получаются белоцветковые растения. Во всех остальных генотипах присутствуют оба доминантных гена, что определяет образование пропигмента и пигмента. Получаются красноцветковые растения. Рассмотренный пример - наиболее простой случай дополнительности генов, когда две группы генотипов - АА вв и аа ВВ - фенотипически неотличимы, т. е. когда действие каждого из взаимодействующих генов в отдельности не проявляется (белая окраска цветков), и это дает расщепление в отношении 9 : 7. По такому типу наследуются содержание цианида в растениях клевера, окраска зерен у некоторых форм кукурузы, устойчивость к ряду ржавчинных грибов у мягкой пшеницы и т. д. Но известны другие, более сложные случаи, когда один или оба дополнительных гена проявляются самостоятельно. Следовательно, характер расщепления во втором поколении будет другим, может быть соотношение 9 : 3 : 4 (наследование окраски шерсти у мышей, окраски луковиц у лука) или 9 : 6 : 7 (наследование формы плода у тыквы). Рассмотрим наследование формы плода у тыквы. При скрещивании плодов сферической формы в первом поколении получаются плоды сферической формы. Появление в F1 дисковидной формы плода тыквы объясняется тем, что в зиготе содержится 2 доминантных гена из взаимодействующей неаллельной пары. Во втором поколении гибридов F2 происходит расщепление в соотношении 9 : 6 : 1. 9 (А - В) : 3 (А - вв) : 3 (а - ВВ) : 1 (аа вв) - дисковидная, сферическая, удлиненная. Дисковидная форма плода возникает в результате взаимодействия двух доминантных генов (АВ), а удлиненная форма - вследствие сочетания их рецессивных аллелей (аа вв). Данный пример комплементарного взаимодействия генов отличается от предыдущего тем, что в случае скрещивания двух форм душистого горошка комплементарное взаимодействие наблюдается только между доминантными неаллельными генами (появление красноцветных растений). А в примере наследования формы плода тыквы, кроме взаимодействия доминантных генов, происходит также взаимодействие рецессивных неаллельных генов, но только в том случае, когда в генотипе отсутствуют доминантные гены обеих неаллельных пар. Это взаимодействие обусловливает появление у 1/16 части всех плодов в F2 удлиненной формы. В обоих рассмотренных случаях при соединении дополнительных генов возникают признаки, свойственные диким формам (красная окраска цветков у гороха, дисковидная форма плода у тыквы). Возможно, это объясняется тем, что в процессе селекции дополнительные доминантные гены разобщались. При скрещивании таких разобщенных отбором генотипов дополнительные доминантные гены вновь соединяются, определяя развитие признаков диких форм. Например, генотип дикого предка Аа Вв в процессе селекции мог быть разложен на генотипы АА вв и аа ВВ. При скрещивании особей с такими генотипами происходит реверсия - вследствие воссоединения в генотипе комплементарных генов. Подобное явление наблюдается при скрещивании двух форм белого горошка, в результате чего образуются растения с дикой красной окраской цветов, или при скрещивании двух рас тыквы со сферической формой плодов происходит возврат к их дикой дисковидной форме.
2.2. ЭПИСТАТИЧЕСКОЕ ВЗАИМОДЕЙСТВИЕ Эпистазом называют подавление (ингибирование) действия одной аллельной пары генов геном другой, неаллельной пары. Ген-подавитель действует на подавляемый ген по принципу, близкому к доминантности, - рецессивности. Разница состоит в том, что эти гены не являются аллельными, т. е. занимают различные локусы в гомологичных или негомологичных хромосомах. Различают доминантный и рецессивный эпистаз. Если обычное аллельное доминирование можно представить в виде формулы А « а, то явление эпистаза выразится формулой А « В (доминантный эпистаз) или а « В (рецессивный эпистаз), когда доминантный или рецессивный ген не допускает проявления другой аллельной пары. Гены, подавляющие действие других, неаллельных им генов, называются эпистатичными, а подавляемые - гипостатичными. Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию. При эпистазе фермент, образующийся под контролем одного гена, полностью подавляет или нейтрализует действие фермента, контролируемого другим геном. Рассмотрим эпистатическое взаимодействие генов на примере наследования окраски зерна у овса. У этой культуры установлены гены, определяющие черную и серую окраску. Оба гена являются доминантными: ген А определяет черную окраску, ген В - серую. В первом поколении в генотипе растений содержатся доминантные гены как черной окраски А, так и серой окраски В. Так как ген А эпистатичен по отношению к гену В, он не дает ему проявиться, поэтому все гибриды в F1 будут черносеменными. В F2 происходит расщепление в соотношении 12 черных : 3 серых : 1 белый: 9 (А - В) : 3 (А - вв) : 3 (а - ВВ) : 1 (аа вв) В девяти типах зигот присутствуют оба доминантных гена А и В, но ген черной окраски А подавляет ген серой - В, вследствие чего семена имеют черную окраску. В трех сочетаниях (АА вв, Аа вв, Аа вв) присутствует только один доминантный ген А, что также обуславливает развитие черносеменных растений. Эта группа по фенотипу совершенно сходна с первой, поэтому 12 растений из 16 будут иметь черные семена. В трех сочетаниях (аа ВВ, аа Вв, аа Вв) присутствует только доминантный ген В, следовательно, все растения с такими генотипами будут серыми. Один генотип (аа вв) представляет собой новую комбинацию, в которой проявится белая окраска зерна, т. к. отсутствуют оба доминантных гена. Этот тип взаимодействия неаллельных генов называют доминантным эпистазом. В отличие от него, при рецессивном эпистазе рецессивная аллель одного гена, будучи в гомозиготном состоянии, подавляет проявление доминантной или рецессивной аллели другого гена. В этом случае вместо ожидаемого при дигибридном скрещивании расщепления (по Менделю) 9 : 3 : 3 : 1, получается соотношение. При эпистазе взаимодействие генов существенно отличается от того, какое мы наблюдаем в явлениях комплементарности. В первом случае проявление одного из генов, влияющих на развитие определенного органа, подавляет проявление другого гена, вследствие чего в потомстве проявляются признаки, свойственные родительским формам. При комплементарности, наоборот, признаки возникают в результате взаимодействия двух неаллельных пар генов.
18. Генетический полиморфизм
сосуществование в пределах популяции (См. Популяция) двух или нескольких различных наследственных форм, находящихся в динамическом равновесии в течение нескольких и даже многих поколений. Чаще всего Г. п. обусловливается либо варьирующими давлениями и векторами (направленностью) отбора в различных условиях (например, в разные сезоны), либо повышенной относительной жизнеспособностью гетерозигот (См. Гетерозигота). Один из видов Г. п. — сбалансированный Г. п. — характеризуется постоянным оптимальным соотношением полиморфных форм, отклонение от которого оказывается неблагоприятным для вида, и автоматически регулируется (устанавливается оптимальное соотношение форм). В состоянии сбалансированного Г. п. у человека и животных находится большинство генов. Различают несколько форм Г. п., анализ которых позволяет определять действие отбора в природных популяциях.
Инбри́динг (от англ. in — «внутри» и breeding — «разведение») — скрещивание близкородственных форм в пределах одной популяции организмов (животных или растений). Термин «инбридинг» обычно используется в отношении животных, а для растений более распространён термин «инцухт» (нем. Inzucht); этот термин также часто используется при описании взаимоотношений между людьми — например, в биографиях.
Инцест является ярко выраженной формой инбридинга, когда скрещивание происходит между особями, связанными прямым родством. Предельная форма инбридинга — самооплодотворение, когда организм оплодотворяет сам себя.
Инбридинг широко используется селекционерами для усиления целевых характеристик породы или сорта. Наиболее распространённая разновидность инбридинга, которая используется при селекции, называется лайнбридингом (англ. linebreeding). При лайнбридинге потомки спариваются с каким-либо своим предком.
Как известно, диплоидный организм получает каждый ген в двух экземплярах (аллелях) — от отца и от матери. Если эти гены различаются, то особь называется гетерозиготной (по данному гену), а если не различаются, то гомозиготной. При инбридинге родители являются родственниками и поэтому имеют много одинаковых генов, в результате чего гомозиготность увеличивается с каждым поколением.
Инбридинг приводит к повышению постоянства фенотипических признаков в потомстве и, в конечном итоге, производится для получения линий генетически идентичных особей (инбредные линии), на которых удобно проводить биологические имедицинские эксперименты.
При близкородственном скрещивании (или самоопылении у растений) может возникать депрессия: уменьшение урожайности растительных культур, измельчание животных, возникновение аномалий и уродств. Это объясняется гомозиготностью по вредным рецессивным генам.
Ассортативность - явление, при котором образование супружеских пар или скрещивание в отношении какого-либо признака происходит не случайным образом: имеется половое предпочтение. Например, в человеческих популяциях отмечается ассортативность по росту. Ассортативность положительна, если пары подбираются по принципу сходства, и отрицательна, если по принципу противоположности.