

5.2.9. Отдел euglenozoa – евгленовые
Отдел назва по типовому роду - Euglena (от греч. еu – хорошо развитый, glene – зрачок, глаз). Объединяет одиночных монадных или амебоидных представителей. Изредка встречаются колониальные формы. Все клетки имеют по 2 базальных тельца с отходящими от них одним или двумя жгутиками. Жгутики по своему строению сходны с таковыми у динофитовых. Евгленовые имеют, как правило, удлиненную, овальную или эллипсоидную форму тела, хотя встречается и шаровидная.
Строение клетки. Клетка у евгленовых водорослей эукариотная и имеет оформленные органеллы с собственными мембранами: ядро, митохондрии, пластиды, комплекс Гольджи, эндоплазматическая сеть.
Форма тела евглен бывает самая разнообразная: веретеновидная, реже яйцевидная, эллипсоидная, клиновидная, цилиндрическая, шаровидная, листовидная, игловидная. Передний конец тела более или менее закруглен, задний может быть вытянутым и заканчиваться заостренным отростком. Клетки могут быть спирально скручены.
Видимых жгутиков может быть 1, 2, 3, 4 и 7. Жгутики отходят от колбообразного впячивания на переднем конце клетки – глотки. Чаще всего у евгленовых два неравных по длине жгутика: один направлен вперед (передний, спинной), другой – назад (задний, брюшной). У некоторых родов, например Евглена, короткий жгутик не выходит за пределы глотки. На поперечном разрезе жгутика виден параксиальный тяж, состоящий из белка. Из-за наличия этого тяжа жгутики евгленовых толще, поэтому они заметнее, чем жгутики других эукариотных водорослей. Жгутики евгленовых покрыты рядом тонких волосков, которые отличаются по своему строению от волосков гетероконтных водорослей (рис. 68).
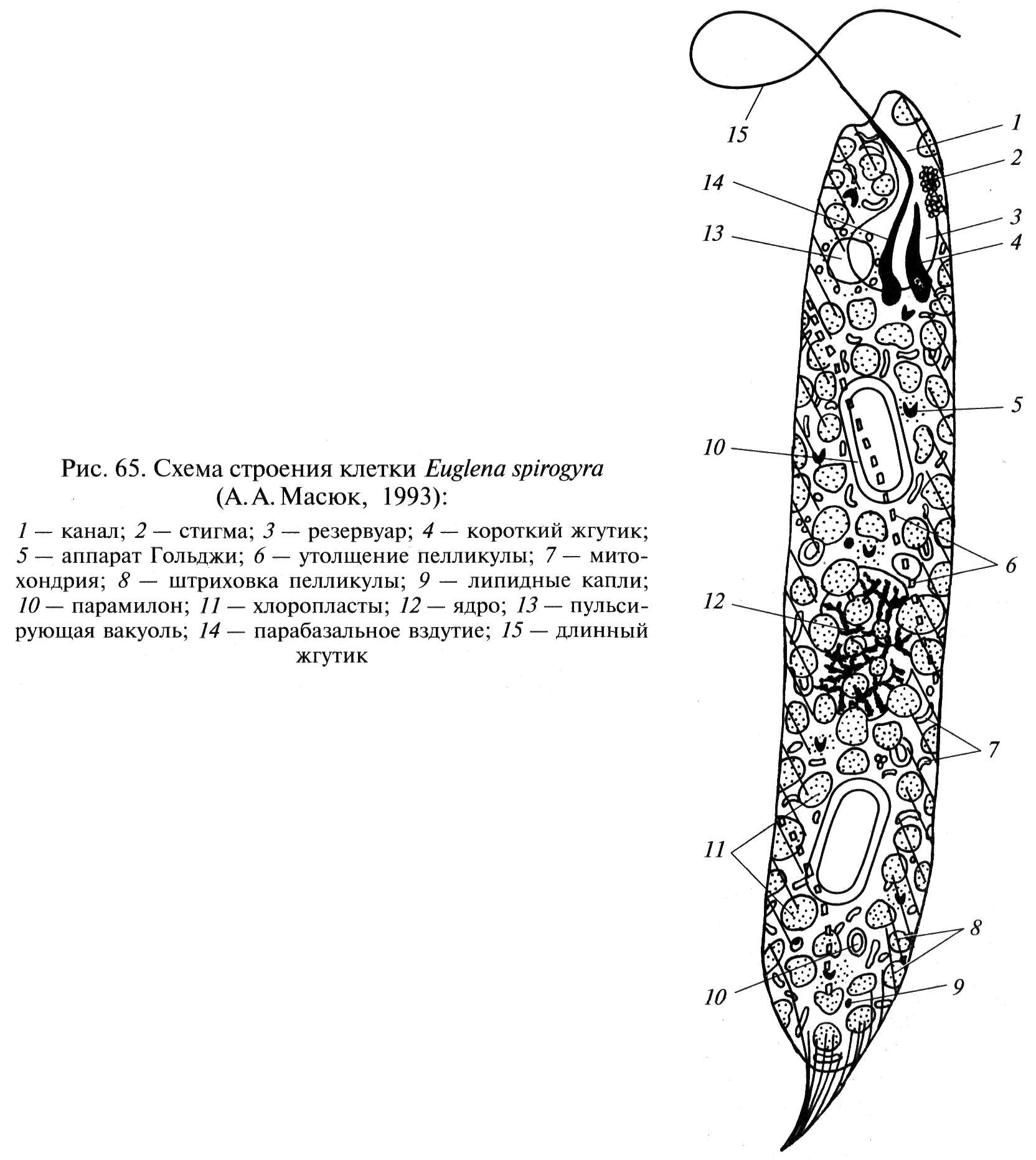
Рис. 68. Схема строения клетки Euqlena spirogyra (по: ): 1 – канал, 2 – стигма, 3 – резервуар, 4 – короткий жгутик, 5 – аппарат Гольджи, 6 –утолщение пелликулы, 7 – митохондрия, 8 – штриховка пелликулы, 9 – липидные капли, 10 – парамилон, 11 – хлоропласты, 12 – ядро, 13 – пульсирующая вакуоль, 14 – парабазальное вздутие. 15 – длинный жгутик
Перипласт имеет в большинстве случаев на своей поверхности штрихи и другие образования, расположенные спиральными рядами от переднего конца к заднему. У некоторых видов евгленовых есть плотные панцири, пропитанные солями железа и марганца и окрашенные в бурый, золотистый или коричневый цвет (рис. 69). Домики-панцири орнаментированы и снабжены отверстиями с горлышками, из которого выходят жгутики.

Рис. 69. Домики эвгленовых водорослей из рода Трахеломонас.
В передней части клетки располагается красное пятно – стигма, которая выполняет роль светочувствительного органа.
У евгленовых оболочка клетки называется пелликулой. Она состоит из цитоплазматической мембраны, расположенного под ней белкового слоя, непрерывного или в виде спиральных подвижно соединенных полос – перипласта, системы микротрубочек и трубчатых цистерн эндоплазматической сети. По химическому составу пелликула на 70–80% состоит из белков, а также содержит липиды и углеводы. Ниже пелликулы располагаются выделительные органеллы – слизистые тельца и мукоцисты. Большинство евгленовых содержат слизистые тельца, имеющие вид мешков, заполненных аморфными или фибриллярными водорастворимыми мукополисахаридами. Они находятся под местами соединения пелликулярных белковых полос и выделяют слизь наружу через свои узкие каналы, расположенные между полосами и открывающимиея порами. Слизистые тельца принимают участие в формировании ножки, образовании домиков, образовании цист, а также в движении.
Хромосомы при окрашивании хорошо видны даже в интерфазе, так как находятся, как и у динофитовых водорослей, в конденсированном состоянии.
Клетки евгленовых имеют одно крупное ядро. Митоз закрытый, веретено деления внутреннее, центриоли отсутствуют. Ядрышко не исчезает во время митоза, в метафазе оно становится значительно удлиненным. Хромосомы формируют неправильную метафазную пластинку. Во время телофазы ядро приобретает форму гантели, становясь сильно удлиненным в анафазе. Оболочки дочерних ядер образуются путем перешнуровки оболочки материнского ядра. Цитокинез (деление цитоплазмы) происходит вслед за делением ядра путем впячивания мембраны по продольной оси клетки.
Форма хлоропласта у разных видов евгленовых широко варьирует: звездчатая, пластинчатая, лентовидная, дисковидная, с пиреноидом или без него. Обычно в клетке содержится несколько хлоропластов. Евглены, растущие при высокой температуре, ультрафиолетовом облучении или на среде со стрептомицином и другими антибиотиками, теряют свои пластиды. Полагают, что хлоропласт евгленовых – это результат вторичного эндосимбиоза бесцветного эукариотного евгленового предка с зеленой эукариотной водорослью. В этом случае 2 его внутренние мембраны объясняются как мембраны хлоропласта зеленой водоросли, а 3-я, наружная, – как мембрана пищеварительной вакуоли клетки-хозяина.
Сократительная (пульсирующая) вакуоль расположена на переднем конце клетки и, возможно, связана с осморегуляторной функцией. Заполненная сократительная вакуоль имеет сферическую форму и окружена рядом мелких добавочных вакуолей. Она изливает свое содержимое в резервуар.
Митохондрии могут сливаться, образуя сеть. Для них характерно наличие уплощенных дисковидных крист. У видов, которые обитают в анаэробных условиях, митохондрии отсутствуют. У евгленовых лизин синтезируется таким же способом, как у настоящих грибов и животных, что является принципиальным отличием евгленовых от всех других фотосинтезирующих организмов.
Общая характеристика.
Евгленовые обладают некоторыми специфическими особенностями: хлоропласты содержат хлорофилл a и b; наряду с этим в фотосинтезе участвуют кератины и ксантофиллы. Однако в клетках евгленовых есть еще один пигмент – астаксантин, который не принимает участия в процессе фотосинтеза. Он сосредоточен в центральной части клетки и не влияет на ее зеленую окраску. При резком повышении освещенности астаксантин перемещается в периферической части клетки и затеняет хлоропласты, окрашивая клетку в красный цвет. Биологический смысл этого явления заключается в регуляции интенсивности падающего на хлоропласт света.
Евгленовые водоросли обладают одной уникальной способностью – им свойственны все три типа питания: автотрофный, гетеротрофный (т. е. питаются готовыми органическими веществами) и голозойный. Последний свойствен видам, которые обладают способностью заглатывать частицы органического вещества и даже некоторые мелкие организмы, что сближает их с животными. У некоторых представителей евгленовых отмечают смешанное питание.
Питательные вещества – парамилон – накапливаются в виде крупных зерен, так называемых парамиллий.
Размножение. Евгленовые водоросли размножаются делением клетки пополам, случается это обычно вечером или рано утром. Иногда перед делением клетки останавливаются, ослизняются, теряют жгутики и после этого наступает процесс деления. Борозда деления проходит от переднего конца клетки к заднему (рис. 70).
При стрессе у евгленовых могут формироваться толстостенные цисты. Образование их связано с потерей жгутиков, увеличением числа парамилоновых гранул, вздутием и округлением клетки, а также с увеличением числа слизистых телец и образованием слоистой слизистой стенки. Образование цист стимулируется низким уровнем питательных веществ и низким соотношением азота и фосфора в среде обитания.

У подавляющего большинства видов евгленовых водорослей половое размножение не известно.
Систематика
В большинстве ботанических классификаций их относят к отделу Евгленовые с единственным классом Евгленовые водоросли – Euglenophyceae, который включает до 1000 видов из 2 порядков: Eutreptiales и Euglenales.
Класс Euglenophyceae – Евгленовые водоросли
Порядок Евтрепциевые – Eutreptiales. Объединяет фототрофных представителей с двумя равными или почти равными жгутиками. Включает роды Евтрепция, Евтрепциелла, Астазия, Дистигма, Рабдонема, Петаломонас, Циклидиопсис и другие.
Род Евтрепция – клетки с глазком и двумя почти равными жгутиками. Многочисленные пластинчатые хлоропласты собраны в звездчатые группы по 25–30 штук. В этих группах они расходятся радиально от центрального пиреноида. Штрихи на пелликуле расположены по спирали. Известно пальмеллоидное состояние и покоящиеся споры. Делится как в подвижном, так и в неподвижном состоянии. Встречается в пресных и морских водах, может вызывать цветение воды.
Род Астазия – клетки имеют один жгутик и многочисленные парамилоновые гранулы. Хлоропласты и глазок отсутствуют. При плавании клетки меняют форму тела. Пелликула спирально заштрихованная. Питание осмотрофное. Вероятно, произошли от окрашенных представителей. Часто встречаются в болотах и водоемах с гниющими растительными остатками
Порядок Евгленовые – Euglenales. Фототрофные и произошедшие от них гетеротрофные представители с осмотрофным и фаготрофным способами поглощения питательных веществ, с двумя неравными жгутиками. У ряда видов имеется специальный аппарат для захватывания и поглощения пищи. Порядок включает роды Евглена, Трахеломонас, Евгленопсис, Гетеронема, Перанема и другие.
Род Евглена (рис. 68) – одноклеточные организмы с одним видимым жгутиком. Клетки уплощенные, вытянутые в длину, иногда спирально закрученные. Пластиды имеют разнообразную форму, с пиреноидами или без. Питание преимущественно миксотрофное. Глазок имеется. Размножаются в подвижном или неподвижном состоянии. Могут образовываться цисты, известно пальмеллевидное состояние. в клетках может находиться большое число гранул с каротиноидом астаксантином. Они маскируют зеленый хлорофилл, придавая клетке красную окраску. при массовом развитии такие виды вызываеют на свету красное "цветение" воды.
Род Перанема – бесцветные пресноводные фаготрофы. Один жгутик направлен вперед, у него подвижен только кончик, другой короткий, расположен в специальной борозде и тянется вдоль клетки. Глазок отсутствует. Питаются другими водорослями, клетками дрожжей. Имеется специальный аппарат для переваривания пищи.
Род Трахеломонас – имеет зеленые метаболичные клетки, расположенные внутри окрашенных домиков. Домик может становиться желтым, коричневым или почти черным. Домик гладкий или орнаментированный (рис. 69). Жгутик выходит через отверстие в домике. При размножении через это отверстие выходит одна или обе дочерние особи, которые вырабатывают в воде собственные домики.
Род Факус – фототрофы с уплощенным неметаболизирующим телом, на заднем конце часто заканчивающимся бесцветным прямым или согнутым отростком. Штрихи на пелликуле расположены продольно или по спирали. Жгутик один, хлоропласты мелкие, многочисленные. Встречаются в различных пресных водоемах.
Экология и значение
Евгленовые водоросли обитают в основном в пресных водах, предпочитая водоемы с замедленным стоком и богатым содержанием органических веществ. Их можно обнаружить в прибрежье озер и рек, в мелких водоемах, включая лужи, на рисовых полях, на сырой почве. В почвах бесцветные представители встречаются на глубине 8–25 см. Окрашенные евгленовые могут вызывать цветение воды, образуя на поверхности воды пленку зеленого или красного цвета.
В значительной степени евгленовые водоросли реагируют на степень минерализации воды: чем она выше, тем беднее их качественный и количественный состав. Некоторые выдерживают высокую соленость воды. По-видимому, недооценивается роль евгленовых в морских местообитаниях. Например, в прибрежных водах Норвегии они могут формировать крупные популяции после весеннего цветения, вызванного диатомовыми водорослями, а у северного побережья Ирландии летом концентрация евгленовых водорослей рода Евтрептиелла достигает 1 млн. на литр.
Евгленовые могут развиваться при широкой амплитуде колебаний рН: 4,5–8,8, однако, виды рода Евтрептиелла, например, могут обитать при рН 3 и даже ниже. Евгленовые способны развиваться в условиях значительного колебания температур (от 0 до 30 °С), но максимум их развития наблюдается летом и осенью. Среди них встречаются холодолюбивые виды, которые могут вмерзать в лед, не теряя жизнеспособности. Большинство евгленовых – свободноживущие представители, но в то же время виды рода Колациум – эпибионты на беспозвоночных животных или эпифиты на нитчатых водорослях. Некоторые евгленовые паразитируют в кишечнике коловраток, нематод, олигохет, плоских червей, амфибий, на жабрах рыб.
Евгленовые – существенный компонент биоценозов различных типов водоемов. Они активно участвуют в круговороте железа, органических кислот. Ими питаются личинки некоторых беспозвоночных. Миксотрофия обеспечивает их участие в процессах самоочищения природных вод и очистке сточных вод. В санитарной гидробиологии некоторые виды используют как биологические индикаторы для проведения биологического анализа состояния органического загрязнения пресных вод.
Евгленовые распространены в пресноводных континентальных водоемах, хорошо прогреваемых, со стоячей водой, богатой органикой, среди зарослей высших водных растений. Богаты ими лужи, канавы, пруды, особенно хорошо удобряемые рыборазводные водоемы, где они часто вызывают «цветение» воды.
Роль евгленовых в природе огромна: они способны к поглощению органических веществ, загрязняющих воду. Активное участие евгленовых в процессах самоочищения воды, особенно в сточных резервуарах, известно давно. Некоторые виды являются биологическими индикаторами санитарного состояния водоемов. Многие виды евгленовых хорошо культивируются, их применяют в экспериментальной биологии в качестве объекта исследований: они служат эталоном для выяснения воздействия на живой организм различных температур, антибиотиков, витаминов, гербицидов и т. д.
Контрольные вопросы
Назовите характерные признаки евгленовых водорослей.
Каковы особенности строения клетки евгленовых водорослей?
Какие пигменты и типы питания известны у евгленовых?
Назовите запасающие вещества, откладывающиеся в клетках евгленовых водорослей.
Каковы особенности размножения евгленовых водорослей?
С какими организмами наиболее сходны евгленовые?
Систематика и типичные представители евгленовых
В каких местообитаниях встречаются евгленовые?
Значение евгленовых для природных экосистем.
Хозяйственное значение евгленовых водорослей.
ЛИТЕРАТУРА
Адрианов А.В., Кусакин О.Г. Таксономический каталог биоты залива Петра Великого Японского моря. Владивосток: Дальнаука, 1998. 349 с.
Алексеев В.В., Лямин М.Я. Биотехнические аспекты фотосинтетического преобразования солнечной энергии // Научные доклады высшей школы. Биол. науки. 1985. № 12. С. 40-45.
Белякова Г.А, Дъяков Ю.Т., Тарасов К.Л. Ботаника: в 4 т. М.: Издательский центр "Академия", 2006. 1280 c.
Великанов Л.Л., Гарибова Л.В., Горбунова Н.П. и др. Курс низших растений. М.: Высш. шк., 1981. 408 с.
Виноградова К.Л. Определитель водорослей дальневосточных морей СССР. Зеленые водоросли. Л.: Наука, 1979. 147 с.
Виноградова К.Л. современные классификации зеленых водорослей (Chlorophyta) и опыт построения их филогенетической системы // Ботан. журн. 1982. Т. 67, № 12. С. 1600-1608.
Виноградова К.Л. К истории формирования морской флоры Chlorophyta. Комаровские чтения. Вып. 35. Л.: Наука, 1984. 66 с.
Вассер С.П., Кондратьева Н.В., Масюк Н.П. и др. Водоросли: Справочник. Киев: Наукова думка, 1989. 608 с.
Возжинская В.Б., Камнев А.Н. Эколого-биологические основы культивирования и использования морских донных водорослей. М.: Наука, 1994. 202 с.
Галышева Ю.А., Коженкова С.И. Морские водоросли и беспозвоночные бухты Киевка: Учебное пособие по летней полевой практике студентов. Владивосток: Изд-во Дальневост.ун-та,2006.160 с.
Гарибова Л.В. и др. Водоросли, лишайники и мохообразные СССР. – М.: Мысль, 1978.
Глезер З.И. Таксономическая значимость признаков у диатомовых водорослей в свете разработки новой классификации Bacillariophyta // Ботан. журн. 1983. Т. 68, № 8. С. 993-1002.
Горбунова Н.П. Альгология: Учеб. пособие для вузов по спец. "Ботаника". М.: Высш. шк., 1991. 256 с.
Грант В. Видообразование у растений. М.: Мир, 1984. 528 с.
Громов Ю.В. Ультраструктура синезеленых водорослей. Л.: Наука, 1976. 186 с.
Громов Ю.В. Строение бактерий. Л.: Изд. Ленингр. ун-та,1985. 191c.
Дроздов А.Л. Биология для физиков и химиков. Учебное издание. Владивосток: Изд-во Дальневост. ун-та, 2005. 414 с.
Жизнь растений. Т.3. Водоросли, гибы и лишайники / Под ред. М.М. Голлербаха. М.: Просвещение, 1977. 488 с.
Звягинцев А.Ю. Морское обрастание в северо-западной части Тихого океана. Владивосток: Дальнаука, 2005. 432 с.
Зинова А.Д. Определитель бурых водорослей северных морей СССР, М.; Л.: Изд-во АН СССР, 1953. 224 с.
Зинова А.Д. Определитель красных водорослей северных морей СССР, М.; Л.: Изд-во АН СССР, 1955. 220 с.
Израэль Ю.А. Экология и контроль состояния природной среды. М.: Гидрометеоиздат, 1984. 560 с.
Калугина-Гутник А.А. Фитобентос Черного моря. Киев: Наук. думка, 1975. 245 с.
Кизеветтер И.В., Суховееева М.В., Шмелькова Л.П. Промысловые морские водоросли и травы дальневосточных морей. М.: Легкая и пищевая промышленность, 1981. 112 с.
Клочкова Н.Г. Флора водорослей-макрофитов Татарского пролива (Японское море) и особенности ее формирования. Владивосток: Дальнаука, 1996. 292 с.
Кондратьева Н.В. Мировоззренческая и методологическая функция систематики живых организмов // Методологические проблемы интеграции ботанических наук. Киев: Наук. думка, 1987. С. 69-80.
Курс низших растений / Под ред. М.В. Горленко. М., 1981. 242 с.
Кусакин О.Г., Дроздов А.Л. Филема органического мира. Часть 1. Пролегомены к построению филемы. СПб: Наука, 1994. 272 с.
Кусакин О.Г., Дроздов А.Л. Филема органического мира. Часть 2. Прокариоты и низшие евкариоты. СПб: Наука, 1998. 380 с.
Кусакин О.Г., Иванова М.Б., Цурпало А.П. Список видов животных, растений и грибов литорали дальневосточных морей. Владивосток: Дальнаука, 1997. 168 с.
Масюк Н.П. О типах морфологической структуры тела водорослей и основных направлениях их эволюции // Ботан. журн. 1985. Т.70, № 8. С. 1009-1018.
Перестенко Л.П. Водоросли залива Петра Великого. Л.: Наука, 1980. 232 с.
Перестенко Л.П. Красные водоросли дальневосточных морей России. СПб: изд-во "Ольга", 1994. 331 с.
Пржеменецкая (Макиенко) В.Ф. Гербарий морских водорослей: Учебное пособие. 2-е изд., испр. и доп. Владивосток: Дальнаука, 2003. 116 с.
Пржеменецкая В.Ф. Экологический ликбез: Учебное пособие. Владивосток: Изд-во Дальневост. ун-та, 2003. 60 с.
Рейвн П. , Эверт Р., Айкхорн С. Современная ботаника в 2-х томах: Пер. с англ. - М.: Мир, 1990. 566 с.
Саут Р., Уиттик А. Основы альгологии: Пер. с англ. - М.: Мир, 1990. 597 с.
Селиванова О.Н. Ревизия систематики морских водорослей-макрофитов на основании молекулярно-филогенетических исследований // Чтения памяти академика Олега Григорьевича Кусакина. Вып. 1. Владивосток: Дальнаука, 2008. С. 161-201.
Чиков П.С. Лекарственные растения – путь к здоровью. – М., 1997.
Abbott I.A.,Hollenberg G.J. Marine algae of California. Stanford, 1976. 827 p.
Adey, W.H. & Vassar, J.M. Colonization, succession and growth rate of tropical crustose coralline algae (Rhodophyta, Cryptonemiales) // Phycologia. 1975. V. 14, N 2. P. 55-69.
Bold, H.C. & Wynne, M.J. Introduction to the Algae. Structure and reproduction. 2nd ed. New York: Prentice-Hall, 1985. 720 p.
Dillon, L.S. Ultrastructure, macromolecules and evolution. New York: Plenum Press, 1981. 708 p.
Guiry, M.D. & Guiry, G.M. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. 2009. http://www.algaebase.org. Last updated: 02 July 2009 by M.D. Guiry.
International Code of botanical nomenclature: Adopted by Fourteenth Intern. botan. congress Berlin, July-Aug. 1987. Koenigstein: Koeltz sci. books, 1988. 328 p.
Kinne, O. Marine ecology. Vol. 1, pt. 2. London: Wiley, 1971.
Dixon, P. Biology of the Rhodophyta. 1973.
Harvey, W.N. Algae. In: Flora Hibernica. Mackay J.T., 1836, Dublin.
Melkonian, M. Systematics and evolution in the algae // Progressive Botany, Vol. 44, P. 315-344.
Parker, S.P. Synopsis and Classification of Living Organisms, Vol. 1 and Vol. 2. New York: McGraw-Hill, 1982.
Segawa, S. Collared illustrations of the seaweeds of Japan. Osaka, Japan, 1962. 175 p.
Silva, P.C. Thallobionta. In: Synopsis and Classification of Living Organisms, Vol. 1 (ed. Parker S.P.). New York: McGraw-Hill, 1982. P. 59-60.
Trainor, F.R. Introductory Phycology. New York: Wiley, 1978. 525 p.
Tseng, C.K. Common seaweeds of China. Bejing, China, 1983. 316 p.
World Catalogue of Algae / World Data Center of Microorganisms (WDC). 1987. 146 p.