

5.2.3. Отдел гетероконтные (разножгутиковые) водоросли – heterokontophyta
У всех гетероконтов похоже устроен жгутиковый аппарат. Имеется 2 жгутика, причем на одном из них есть очень характерные трубчатые трехчленные перистые выросты, или волоски – мастигонемы. Именно наличие двух неравных жгутиков послужило основанием для того, чтобы назвать эту обширную группу водных организмов разножгутиковыми водорослями, или гетероконтами. Помимо различий в длине, жгутики различаются и морфологически: главный жгутик состоит из оси и перисто расположенных на ней мерцательных волосков, боковой жгутик гладкий бичевидный. Сходен также состав пигментов – хлорофиллы а и с и многочисленные каротиноиды, а также строение хлоропластов. Хлоропласты окружены четырьмя мембранами, причем две наружные мембраны связаны с эндоплазматической сетью клетки.
В составе отдела Гетероконтные (Разножгутиковые) водоросли объединяют несколько групп водорослей в ранге класов: Бурые, Золотистые, Желтозеленые., Рафидофитовые, Синуровые, Феотамниевые, Евстигмовые, некоторые ранее рассматривались как самостоятельные отделы.
Класс Бурые водоросли – Phaeophyceae (Fucophyceae)
Название класса «бурые водоросли» отражает окраску их таллома (от греч. рhaios – смуглый). Окраска обусловлена наличием большого количества бурых и желтых пигментов – помимо хлорофиллов а и с и β-каротина в их хлоропластах имеется избыток ксантофиллов, особенно фукоксантина.
Многоклеточные, преимущественно крупные морские водоросли, особенно широко распространенные в холодных водах обоих полушарий. Ни одноклеточные, ни колониальные формы среди бурых водорослей неизвестны. Известно около 1,8 тыс. видов.
Строение клетки. Клетка у видов бурых водорослей эукариотная: она имеет оформленные органеллы с собственными мембранами: ядро, митохондрии, пластиды и др. Зеленые пигменты – хлорофиллы а и с – маскируются дополнительными: оранжевыми каротинами и желто-коричневыми ксантофиллами; из последних преобладает фукоксантин. В клетках бурых водорослей встречается от одной до многих пластид. Поверх наружной клеточной мембраны клетки бурых водорослей обычно вырабатывают жесткую клеточную стенку.
Ядро чаще всего одно, но иногда ядер несколько. У некоторых представителей по мере увеличения возраста клетки количество ядер увеличивается. Ядрышко в ядре всегда одно.
Хлоропласты вегетативных клеток бурых водорослей имеют глазок (стигму) и пиреноиды, которых бывает один или несколько; у некоторых видов пиреноиды отсутствуют. Глазок, состоящий из 30–40 липидных глобул, выполняет функцию линзы, фокусирующей свет на жгутиковое вздутие, которое является фоторецептором. Зооспоры и гаметы бурых водорослей обычно не имеют глазка. В пиреноидах откладываются запасные вещества. Хлоропласты чаще всего мелкие, дисковидные, париетальные. Их форма может быть звездчатой, лентовидной или пластинчатой. Форма хлоропластов в клетках бурых водорослей может изменяться по мере старения клетки. Оболочка хлоропласта состоит из четырех мембран, причем две наружные мембраны связаны с ЭПС -эндоплазматической сетью клетки.
Целлюлоза, из которой состоит стенка клеток бурых водорослей – ее иногда называют альгулезой, по свойствам отличается от целлюлозы высших растений. Кнаружи от целлюлозной стенки располагается сильно ослизняющийся пектиновый слой, образованный в основном альгиновой кислотой и ее солями и соединениями с белковыми веществами. Альгиновая кислота также обнаружена только у бурых водорослей.
Общая характеристика.
Все бурые водоросли являются фототрофами. Продукты фотосинтеза - специфический углевод хризоламинарин, спирт маннит и липиды. Липиды накапливаются в виде капель масла. Хризоламинарин откладывается вне хлоропласта: он располагается в специальных вакуолях в цитоплазме около пиреноида. Маннит выполняет резервную, а также и осморегуляторную функцию; его концентрация внутри клетки зависит от изменения солености воды.
В молодых клетках бурых водорослей содержатся мелкие и многочисленные вакуоли, которые с возрастом становятся крупнее за счет слияния. В цитоплазме расположены особые образования – физоды, которые содержат флоротаннины. Эти вещества известны только у видов бурых водорослей. Функция флоротаннинов, возможно, связана с защитой талломов водорослей от выедания животными, например, моллюсками. Флоротаннины бурых водорослей, вероятно, ингибируют поселение на поверхности таллома эпифитных водорослей и животных. Предполагается, что флоротаннины участвуют в защите от радиционного повреждения и в избирательном поглощении из водной среды ионов тяжелых металлов. Бесцветные флоротаннины на воздухе окисляются с образованием пигмента фикофеина, который придает высушенным бурым водорослям характерную темную окраску.
Строение таллома. Среди видов бурых водорослей отсутствуют одноклеточные и колониальные формы, все особи многоклеточные. Форма талломов бурых водорослей самая разнообразная. Это нити, стелющиеся или вертикально стоящие, однорядные или многорядные; массивные сплошные или черепитчато соединенные корки и корочки; тонкие слизистые пластинки или плотные кожистые пластины на стволике, простые или рассеченные; пузыри и мешки; трубки и шнуры; различным образом разветвленные тонкие кустики или мощные кусты. Прикрепление талломов осуществляется с помощью ризоидов или подошвы. Для удержания в вертикальном положении у крупных бентосных форм бурых водорослей образуются воздушные (плавательные) пузыри, заполненные газом (рис. 29).

Рис. 29. Внешний вид бурых водорослей: 1 – Фукус; 2 – Сахарина; 3 – Алария; 4 – Саргассум
Наиболее сложно среди видов бурых водорослей устроены талломы представителей порядков Ламинариевые и Фукусовые. Они имеют признаки тканевой дифференцировки со специализацией клеток таллома (рис. 30). В их слоевищах можно различить кору, состоящую из интенсивно окрашенных клеток и сердцевину, состоящую из бесцветных клеток, которые часто собраны в нити. У Ламинариевых в сердцевине образуются также ситовидные трубки и трубчатые нити. Сердцевина выполняет не только транспортную функцию, но и механическую, так как в ней находятся нити с толстыми продольными стенками. Между корой и сердцевиной у многих бурых водорослей может находиться промежуточный слой из крупных бесцветных клеток.
Рост таллома у представителей бурых водорослей чаще всего интеркалярный (вставочный) и апикальный (верхушечный), реже базальный. Интеркалярный рост может быть диффузный, или имеется зона роста. У крупных форм вставочная (интеркалярная) меристема, или активно делящаяся группа клеток, расположена в месте перехода «черешка» в «листовую» пластинку. Крупные водоросли также имеют меристематическую зону на поверхности таллома, так называемую меристодерму (своеобразный аналог камбия высших растений).
Размножение
у бурых водорослей вегетативное, бесполое
и половое. Гаметы формируются в
многогнездных
гаметангиях, зооспоры –
в

Рис. 30. Внутреннее строении Ламинарии (по: Л.Л. Великанов и др., 1991): А – продольный срез стволика; Б – поперечный срез стволика; В – срез пластины с сорусами зооспорангиев
одногнездных спорангиях.
Вегетативное размножение у ряда видов бурых водорослей может осуществляться участками таллома; у видов рода Сфацелярия – выводковыми веточками (выводковыми почками); у видов рода Фукус - группой клеток на подошве, которые способны к дифференцировке в новый таллом.
Бесполое размножение бурых водорослей обычно происходит с помощью зооспор. Некоторые виды размножаются неподвижными тетраспорами или моноспорами. Споры бесполого размножения формируются в результате мейоза и последующих митозов в одногнездных спорангиях. Половой процесс – изогамия, гетерогамия и оогамия. Гаметы образуются в многогнездных гаметангиях - особых группах клеток. В каждом «гнезде» гаметангия формируется одна гамета.
Для бурых водорослей известны половые феромоны – растворимые вещества, которые координируют активность клеток при размножении. Феромоны, которых известно по крайней мере десять, могут или стимулировать раскрытие антеридиев, или привлекать мужские гаметы к женским. Они также участвуют в изоляции видов. Полагают, что феромоны образуются у большинства видов бурых водорослей, имеющих половой процесс.
Жизненный цикл. У бурых водорослей можно выделить два основных типа жизненных циклов. Один тип – гаплодиплобионтный. Спорофиты диплоидны, т.е. все клетки содержат полный набор хромосом. Споры бесполого размножения формируются в одногнездных спорангиях. При образовании спор происходит редукционное деление – мейоз. Такая редукция называется спорической. Образовавшиеся после мейоза клетки (споры) гаплоидны, т.е. содержат половинный набор хромосом. Гаплоидные зооспоры и тетраспоры прорастают в гаплоидный гаметофит, на котором в многогнездных гаметангиях формируются гаметы. Образовавшаяся после слияния гамет диплоидная зигота прорастает в диплоидный спорофит.
Другой тип – диплобионтный; спорофиты, споры и гаметофиты диплоидны; редукционное деление происходит при образовании гамет. Такая редукция называется гаметической. После слияния гаплоидных гамет образуется диплоидная зигота и прорастает в диплоидный спорофит. Этот тип распространен у видов ламинариевых (рис. 31).
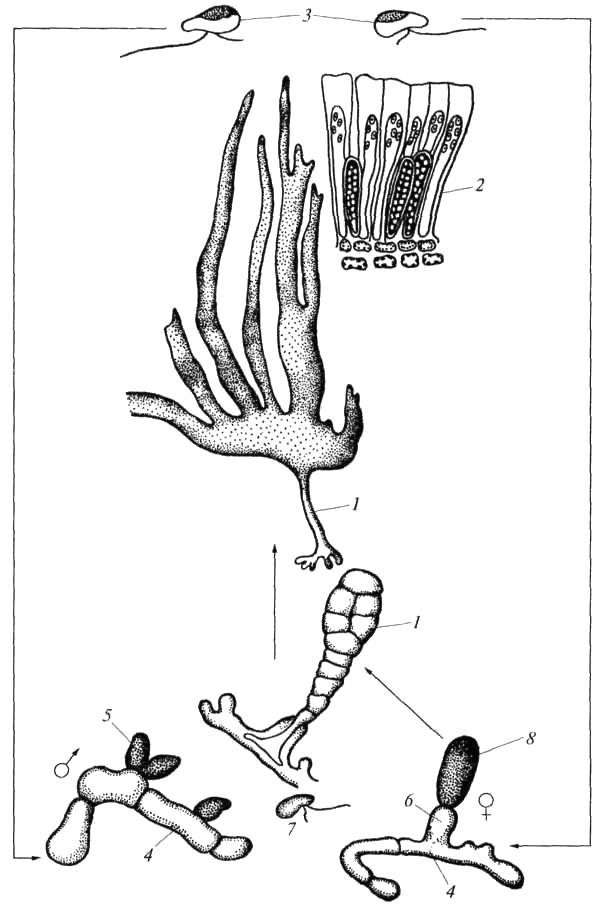
Рис. 31. Жизненный цикл Сахарины (по: Л.Л.Великанов и др., 1991): 1 – спорофит, 2 – сорусы зооспорангиев, 3 – зооспоры, 4 – гаметофит, 5 – антеридий, 6 – оогоний, 7 – антерозоид, 8 –зигота (оплодотворенная яйцеклетка)
На макроскопическом спорофите Сахарины, в верхней части пластины, полосами развиваются сорусы (группы) зооспорангиев. В зооспорангиях после редукционного деления образуются зооспоры с двумя неравными жгутиками. Они вырастают в микроскопические гаметофиты: мужские с антеридиями и женские с оогониями. Оплодотворенная антерозоидам яйцеклетка сразу, без периода покоя, прорастает и образует спорофит. На этом жизненный цикл Сахарины завершается.
Фукусовые водоросли не имеют спорофиов. У Фукуса мужские и женские половые органы – антеридии со сперматозоидами и концептакулы с оогониями и яйцеклетками – находятся в в углублениях слоевища – скафидиях , мужские и женские скафидии могут находиться на одном рецептакуле или в разных рецептакулах. Оогонии сидят на клетке-ножке, возникающей на стенке концептакула (рис. 32, В, Г). Клетка оогония претерпевает три деления
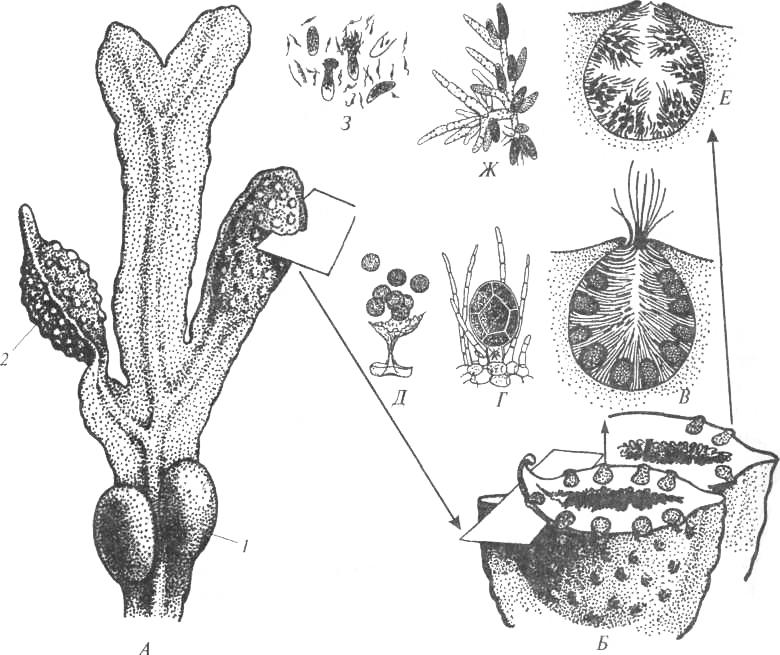
Рис. 32. Размножение Фукуса (по: B.Fott, 1971; W.Braune et al., 1974): A – фрагмент таллома (1 – воздушный пузырь, 2 – рецептакул); Б – разрез через рецептакул; В – разрез женского скафидия; Г – оогоний; Д – выход яйцеклеток; Е – разрез мужского скафидия; Ж – веточка с антеридиями; 3 – выход сперматозоидов
ядра, первое из которых редукционное, в результате чего в оогонии образуется 8 яйцеклеток.
Антеридии обычно формируются на поддерживающих их разветвленных нитях – парафизах (рис. 32, Е, Ж). В результате нескольких делений образуется 64 сперматозоида. Сперматозоиды Фукуса имеют глазок, расположенный внутри редуцированного хлоропласта. В переднем конце клетки сперматозоида 13 микротрубочек формируют хоботок – особую структуру, которая, как предполагают, принимает участие в определении женского полового аттрактанта.
Сперматозоиды привлекаются к яйцеклетке Фукуса неспецифичным феромоном фукосерратеном, выделяемым яйцеклеткой. Узнавание же на видовом уровне связано со специфическими олигосахаридами на яйцеклетке и сперматозоиде. На поверхности яйцеклетки имеются различные участки, каждый из которых содержит особые органические соединения - гликопротеины. Сперматозоиды содержат гликопротеины, организованные в участки на мембране и выростах переднего жгутика, а также на поверхности самого сперматозоида. При встрече с яйцеклеткой сперматозоид движется над оболочкой яйцеклетки, как бы исследуя ее с помощью своего переднего жгутика. Если гликопротеины сперматозоида комплементарны гликопротеинам яйцеклетки, только тогда происходит узнавание сперматозоида яйцеклеткой.
Систематика
Ископаемые находки, которые могут иметь отношение к бурым водорослям, известны из среднего силура (около 425 млн. лет). Но эти находки имеют сходство и с некоторыми современными зелеными и красными водорослями. Ископаемые находки, которые точно можно связать с современными бурыми водорослями, относятся к миоцену (5–25 млн. лет). Эти водоросли напоминают современную Диктиоту. Возраст бурых водорослей, определяемый молекулярными методам, составляет 155–200 млн. лет.
Тип организации таллома, наличие или отсутствие пиреноида, способ роста, тип полового размножения (изогамия, гетерогамия, оогамия) и жизненного цикла используют для выделения порядков бурых водорослей. В последние годы активно пересматривается система бурых водорослей. В разных классификационных схемах в составе Класса Бурые водоросли выделяют от 7 до 14 порядков.
Порядок Десмарестиевые – Desmarestiales. Водоросли кустистые, многократно разветвленные, с вальковатым, плоским или уплощенным слоевищем. Прикрепляется к субстрату хорошо развитой подошвой. На поверхности слоевища развиваются волоски.
Род Десмарестия (рис.33). Бурые или оливковые кусты с хорошо выраженным главным побегом, до 0.8–1 м высоты. Прикрепляется конусовидной подошвой. Ветвление 3–4 порядков, поочередное или супротивное. Ветви последних порядков плоские или, реже, нитевидные. Виды морские, обитают в арктических и умеренных широтах на глубине 1–20 м, образуют заросли или растут единично.

Рис. 33. Десмарестия (р. Desmarestia)
Порядок Ральфсиевые – Ralfsiales. Представители порядка имеют cлоевище в виде корок различного строения либо состоят из базальной корки и вертикальных побегов.
Род Аналипус. Слоевище состоит из многолетней коркообразной базальной части и отходящих от нее однолетних, оливково-бурых, вертикальных побегов высотой до 40 см. Многогнездные спорангии цилиндрические по формы и двурядные по строению. Одногнездные спорангии овальные, развиваются среди ассимиляционных ветвей. Виды рода широко распространены в морях умеренных широт, образуют пояса в приливо-отливной зоне.
Порядок Диктиотовые – Dictyotales. Для видов порядка характерен апикальный рост. В вегетативных клетках имеются многочисленные дисковидные хлоропласты без пиреноидов. Таллом паренхиматозный, характерна изоморфная смена поколений. Половой процесс оогамный, бесполое размножение неподвижными спорами, сперматозоиды имеют только один жгутик. Жгутик длинный, с перистыми выростами и направлен вперед. Представители порядка встречаются в основном в тропических и, реже, в умеренных морях.
Род Диктиота (приложение,4А). Имеет слоевище в виде плоских оливково-бурых дихотомически разветвленных кустиков высотой до 11–15 см. Ветви линейные, узкие, не имеют жилок, с раздвоенными вершинками. Тетраспорангии образуют на поверхности слоевища сорусы, которые имеют неопределенную или кольцеобразную форму. Распространен в морских водах умеренных и теплых широт.
Порядок Ламинариевые – Laminariales. Ламинариевые имеют слоевище, состоящее из пластины, стволика и органов прикрепления – подошвы или ризоидов. Пластина ровная или морщинистая, цельная или рассеченная, с отверстиями, складками, булями или без них, длиной от нескольких десятков сантиметров до 20–40 м. Стволик разветвленный или чаще неразветвленный. Спорофиты однолетние или многолетние, возраст некоторых видов насчитывает 11–18 лет. В коре многих ламинариевых расположены слизистые каналы.
Наиболее популярными представителями бурых водорослей являются ламинарии (виды родов Ламинария или Сахарина), или «морская капуста», которые распространены преимущественно в Тихом океане. Они встречаются от поверхности (уреза) воды до глубины 200 м. Растут ламинарии предпочтительно на глубине 2–20 (30) м, в местах с постоянным движением воды, прикрепляясь к камням и скалам. Ламинариевые образуют водорослевые пояса вдоль побережья морей Дальнего Востока. Наиболее мощные заросли встречаются у Курильских островов и у о. Хоккайдо, где сконцентрировано около половины мировых ресурсов ламинариевых. Ежегодный мировой сбор ламинариевых водорослей достигает 2 млн. тонн сырой массы, при этом более 1 млн. тонн дает производство ее марикультуры в Китае.
Род Сахарина (Ламинария). Слоевище длиной от 0,5 до 5–10 м, имеет толстую блестящую пластину, упругий стволик и густо переплетенные ризоиды, которыми прикрепляется к различным твердым субстратам. Края пластины гладкие или волнистые, на поверхности пластины часто имеются выпуклости и складки. Скопления спорангиев – сорусы – образуются на обеих сторонах пластины, после выхода спор верхняя часть пластины разрушается. Живет от двух до нескольких лет. Сахарины являются ценным промышленным сырьем и объектом марикультуры. Сахарина японская образует заросли в северной половине Японского моря, а также у южного и юго-восточного берега Сахалина и у Южных Курильских островов. Сахарина японская является наиболее ценным промысловым видом, она отличается превосходными вкусовыми качествами и используется для приготовления разнообразных блюд, консервов, кондитерских изделий.
Порядок Сфацеляриевые – Sphacelariales.
Бурые водоросли, относящиеся к этому порядку, имеют ветвящийся таллом с апикальным ростом. Ветви с волосками. Вегетативные клетки имеют многочисленные хлоропласты без пиреноидов. Жизненный цикл у большинства с изоморфной сменой поколений, половой процесс - изогамия или гетерогамия. Водоросли образуют небольшие бурые пучочки до 3 см в высоту и разветвленные кустики до 20 см в высоту на камнях или других водорослях; есть эндофитные представители.
Род Сфацелярия. Имеет тонконитчатое слоевище. Ветвящиеся нити образуют небольшие, до 0,5–3 см высоты, пучочки бурого цвета. Вегетативное размножение осуществляется специализированными, часто трехлучевыми выводковыми веточками - пропагулами. Половой процесс изогамный. Женские гаметы выделяют феромоны эктокарпен и мультифиден. Виды рода распространены в морях как полярных, так и тропических широт.
Порядок Сцитосифоновые – Scytosiphonales. Представители с кишковидным, пузыревидным, пластинчатым или мешковидным, часто полым слоевищем до 25 см длины. Кора однорядная или двурядная, внутренняя часть (стенка) слоевища образована несколькими слоями крупных клеток. На поверхности слоевища развиваются многогнездные спорангии, которые собраны в сорусы, поддерживающие их нити – парафизы и пучки волосков.
Род Сцитосифон представляет собой тонкостенные бурые трубки 4–25 см длины (приложение,4Б). Слоевище прикрепляется небольшой подошвой и имеет редкие поперечные перетяжки. Встречается в прибрежной зоне морей в различных широтах, часто образует пояса в приливо-отливной зоне.
Порядок Фукусовые – Fucales. Имеют многолетнее слоевище в виде крупных неправильно или дихотомически ветвящихся кустов до 1–3 м длины. Прикрепляются к субстрату конусовидной подошвой. Слоевища имеют побеги, филлоиды (листоподобные образования), рецептакулы (вместилища гамет) и плавательные (воздушные) пузыри. Встречаются в приливо-отливной зоне всех умеренных и арктических морей, где образуют мощные заросли – пояса.
Род Саргассум (рис. 34, В). Обильно разветвленные оливково-бурые или коричневые кусты высотой 0,5–1,5 м. Прикрепляются конической подошвой. Филлоиды развиваются на боковых побегах, отходящих от цилиндрического стволика, в нижней и средней частях слоевища. Филлоиды кожистые, с центральным ребром, 0,2–2 см длины. Плавательные пузыри овальные или округлые, с выростами или без них. Саргассы широко распространены в умеренных и субтропических широтах от нижней литорали до глубины 10 м, где образуют плотные или разреженные заросли. В южной части Тихого океана – так называемом Саргассовом море – образуют обширные плавающие скопления, в которых размножаются только вегетативным путем.
Род Фукус (рис.32). Слоевище кожистое, кустистое, ветвится дихотомически, высотой 25–50 см. У побережья дальневосточных морей растет несколько лет, обитает на литорали. Часто образует большие заросли на мелководье. Для видов этого рода характерно дихотомически
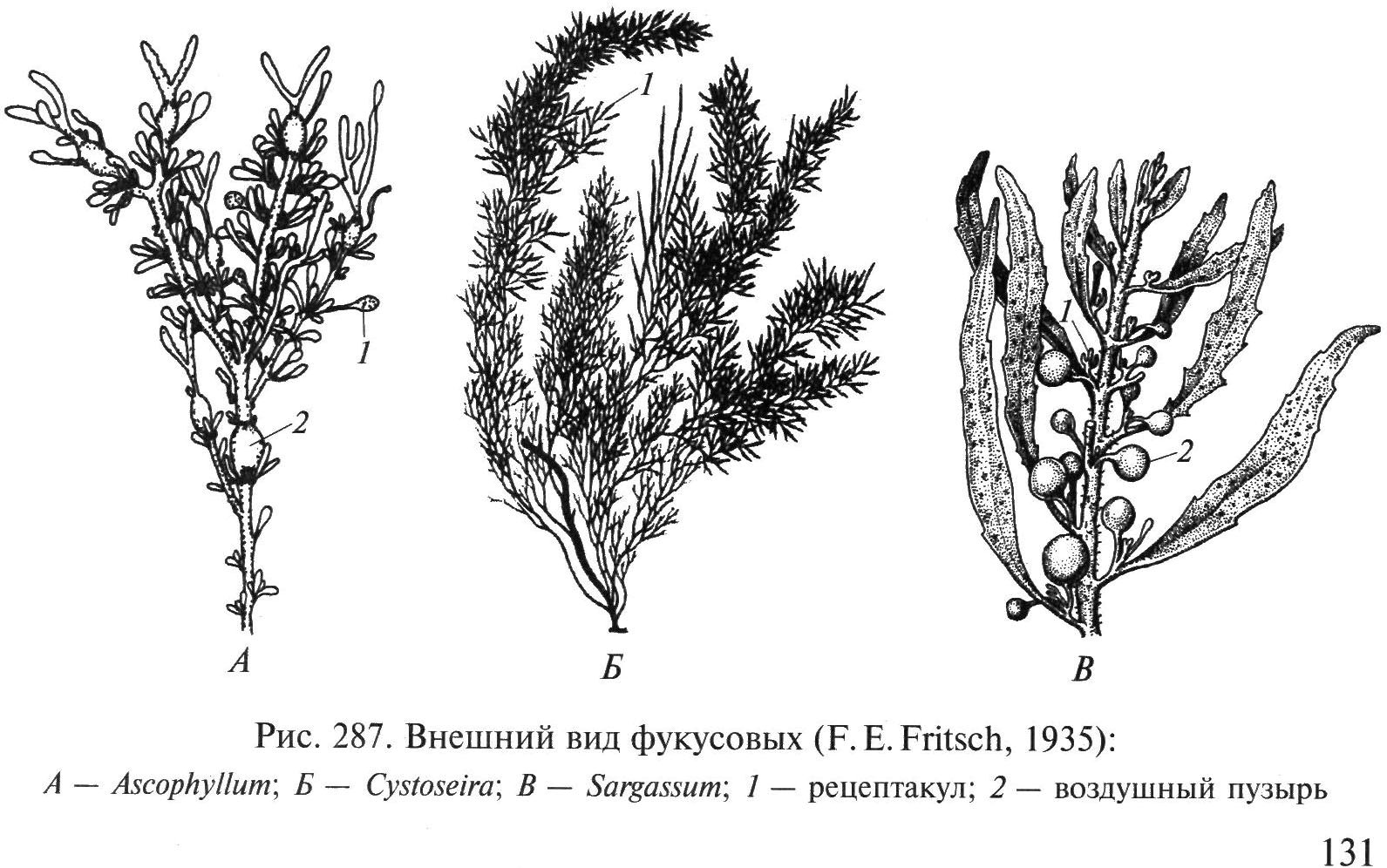
Рис. 34. Внешний вид фукусовых водорослей: А – Аскофиллум; Б – Цистозира; В – Саргассум; 1 – рецептакул, 2 – плавательный пузырь
разветвленное слоевище с плоскими ветвями, имеющими продольное ребро, которое прикрепляется к камням конической подошвой. Фукусы используют в качестве удобрений, как корм для скота, для производства кормовой муки, альгинатов, применяют в медицине для лечения зоба, коррекции веса и т.д.
Порядок Эктокарповые – Ectocarpales. Водоросли, которые сюда относятся, микроскопические, свободноживущие, эпифиты, эпизоиты или эндофиты. Органы размножения представлены одногнездными (спорангии) и многогнездными (гаметангии) спорангиями; они длинноцилиндрические, короткоцилиндрические или стручковидные.
Род Эктокарпус. Слоевище в виде мягких бурые кустиков до 2 см высоты. Кустики образованы однорядными, поочередно, дихотомически или неправильно разветвленными нитями. Многогнездные спорангии стручковидные, образуются сбоку боковых ветвей на 1–3-клеточной ножке. Виды распространены в морях различных широт, в чистых и загрязненных водах, на литорали и в верхней части сублиторали. Эпифиты, встречаются и в обрастании антропогенных субстратов.
Экология
Подавляющее большинство представителей бурых водорослей живут в морских водоемах. В пресных водах встречаются только 8 видов. Встречаются как однолетние, так и многолетние виды, возраст которых может достигать 15–18 лет. Ламинариевые водоросли Нереоцистис и Макроцистис, которые обитают вдоль тихоокеанского побережья Северной Америки, имеют максимальные для всех водорослей размеры – до 30–40 м в длину. Они формируют гигантские подводные леса в морях Дальнего Востока. Прикрепляются бурые водоросли к самым разнообразным субстратам – скалам, камням, гравию, раковинам и панцирям морских животных, другим водорослям. Некоторые мелкие формы бурых водорослей живут внутри тканей других водорослей как эндофиты. Большинство видов бурых живут в прикрепленном состоянии. Оторванные от грунта слоевища сносятся течением в спокойные места с илистым дном и продолжают там существовать. Виды с воздушными пузырями на слоевище, оторвавшись от грунта, всплывают к поверхности, образуя большие плавающие скопления, особенно в районах со стабильным круговым течением, как в Саргассовом море. Они существуют в таких скоплениях довольно долгое время и размножаются только вегетативным способом.
Бурые водоросли широко распространены во всех морях, но наибольшего развития они достигают в морях умеренных и приполярных широт. Их крупные слоевища растут, прикрепившись к скалам и камням, а в спокойных местах у берега и на большой глубине могут удерживаться даже на створках раковин моллюсков и гравии. Встретить их можно на разной глубине – от литоральной зоны, где во время отлива они часами находятся вне воды, до 40–200 м. В районе Гавайских островов на глубине 180 м обнаружены виды рода Саргассум, а Ламинария в Адриатическом море была найдена на глубине 200 м. Самые обильные заросли бурых водорослей наблюдаются на глубине 6–15 м, где наилучшие условия освещения и постоянное движение воды, которое приносит к их слоевищам биогенные вещества и ограничивает поселение растительноядных животных - фитофагов.
Класс Золотистые водоросли – Chrysophyceae
Одноклеточные, колониальные или многоклеточные организмы, обычно имеющие жгутиковую стадию в жизненном цикле. Клетки одноядерные, с одним или несколькими хлоропластами. Стигма обычно имеется (рис. 36). Жгутиков 1–2, неравных. Сократительная вакуоль расположена в передней части клетки. Наружные покровы клеток этих организмов очень разнообразны. У наиболее примитивных видов клетка одета нежным перипластом, позволяющим образовывать выпячивания разной формы (ризоподии, псевдоподии), у других плазмалемму покрывает твердая целлюлозная оболочка, иногда сильно ослизняющаяся. Домики у золотистых водорослей бывают разной формы: вазообразные, шаровидные, яйцевидные, цилиндрические; с одним или несколькими отверстиями. Клетка прикрепляется к основанию домика с помощью гибкой ножки или может быть свободной. У некоторых хризофит на поверхности клетки откладываются известковые образования – кокколиты – различных форм и размеров, а у кремниевых жгутиконосцев имеется внутренний кремниевый скелет. Разнообразие строения этих скелетов позволяет использовать остатки тех или иных форм ископаемых силикофлагеллят для определения возраста содержащих их геологических пород. У более высокоразвитых представителей отдела золотистых водорослей клетки одеты панцирем, состоящим из кремниевых чешуек (иногда несущих шипы), или заключены в домики, через отверстия которых наружу выходят жгутики или псевдоподии.
Домики у видов золотистых водорослей вазообразные, шаровидные, яйцевидные, цилиндрические, с одним или несколькими отверстиями. Клетка прикрепляется к основанию домика с помощью гибкой ножки или может быть свободной.
Пигменты – хлорофиллы а и с, фукоксантин. Для хризофит характерно наиболее высокое среди гетероконтов содержание фукоксантина (55-93% от всех каротиноидов).
Большинство видов - пресноводные фототрофы, но встречаются гетеротрофы и фаготрофы.
Запасные вещества – хризоламинарин и масла.
Среди одноклеточных форм золотистых водорослей преобладают пресноводные планктонные виды, а среди многоклеточных – донные или эпифитные. Встречаются в основном в прохладных морский и пресных водоемах. Известно около 360 видов.

А В Г
Рис. 36. Золотистая водоросль (по: С. Hoek van den et al., 1995): А – строение клетки; Б – корешковая система; В – трехчастная мастигонема; Г – срез части хлоропласта; 1 – базальное тело; 2 – базальное вздутие; 3 – короткий жгутик; 4 – стигма; 5 – ризопласт; 6 – ядро; 7 – ядерная мембрана; 8 – ядрышко; 9 – митохондрия; 10 – вакуоли; 11 – слизь; 12 – плазмалемма; 13 – везикулы; 14 – слизистое тело; 15 – липиды; 16 – хлоропласт; 17 – аппарат Гольджи; 18 – сократительная вакуоль; 19 – длинный жгутик: 20 – 1-й корешок; 21 – 2-й корешок; 22 – 3-й корешок; 23 – 4-й корешок; 24 – хлоропластный нуклеоид; 25 – ХЭС; 26 – оболочка хлоропласта; 27 – опоясывающая ламелла; 28 – ламелла; 29 – длинный латеральный филамент; 30 – короткий латеральный филамент; 31 – терминальный филамент; 32 – трубчатая часть мастигонемы
Структура таллома у золотистых водорослей чаще всего монадная, но может быть очень разнообразной: коккоидной, пальмеллоидной, плазмоидальной, амебоидной, нитчатой, паренхиматозной (рис. 37).
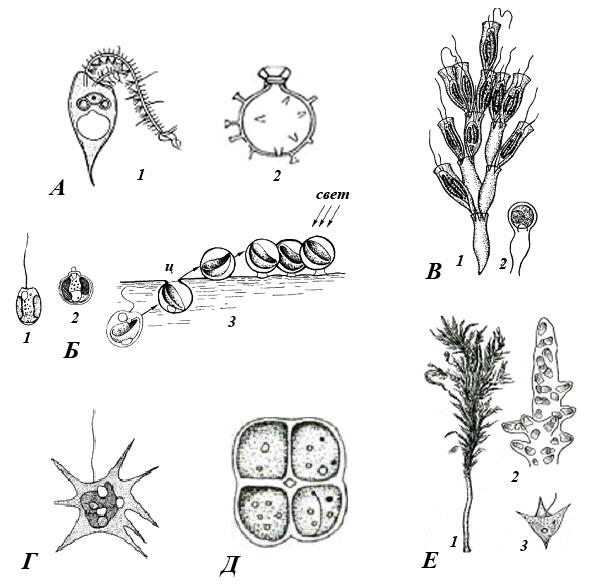
Рис. 37. Золотистые водоросли (по: Л.Л. Великанов и др., 1981): А – Охромонас: 1 – внешний вид, 2 – циста; Б – Хромулина: 1 – внешний вид, 2 – циста, 3 – схема образования пленки из цист; В – Динобрион: 1 – общий вид колонии, 2 – циста; Г – Хризамеба; Д – Гиббердия; Е – Гидрурус: 1 – общий вид колонии, 2 – кончик ветви, 3 – зооспора
Размножение. У золотистых водорослей встречаются три способа размножения: вегетативное, бесполое и половое.
Вегетативное размножение происходит путем продольного деления клетки или распада колонии на части за счет деления клеток в одном, двух или трех направлениях.
Половой процесс (клетки золотистых водорослей диплоидны) – изогамия или конъюгация. В результате полового процесса, а также в неблагоприятных условиях, образуются цисты с толстой оболочкой, содержащей кремний.
Бесполое размножение осуществляется одно- или двужгутиковыми зооспорами, которые развиваются в обычных вегетативных клетках или зооспорангиях. Половой процесс: голо-, изо- и автогамия. Гологамия – вид полового процесса, при котором две сходные с вегетативными клетки сливаются на передних концах в двуядерную зиготу, которая превращается затем в окремнелую цисту.
Систематика
Из Золотистых водорослей в конце прошлого столетия на основании результатов молекулярно-генетических исследований выделен класс Синуровые. Также выделен класс Феотамниевые – из Желтозеленых и Золотистых водорослей. Поэтому объем класса Chrysophyceae сократился и в настоящее время насчитывает около 360 видов, относимых к трем порядкам: Chromulinales, Hibberdiales и Hydrurales. Форма, строение и количество жгутиков, а также пигментный состав и структура тела являются важными систематическими признаками.
Порядок Хромулиновые – Chromulinales. К этому порядку относят организмы с монадным, пальмеллоидным и амебоидным типами дифференциации таллома. Монадные клетки с одним жгутиком, видимым в световой микроскоп.
Род Хризамеба (рис. 37, А) – пресноводные амебоидные водоросли, они имеют в жизненном цикле жгутиковую стадию. Зигота превращается в стоматоцисту.
Род Хромулина (рис. 37, В) - одноклеточные свободноплавающие организмы, обитающие преимущественно в пресных водах. Клетки голые; имеется одна или две золотистые пластиды.
Род Динобрион (рис. 37, Г) – одноклеточные и колониальные, свободноплавающие или прикрепленные представители. Монады находятся в вазообразных домиках, образованных целлюлозными микрофибриллами. Помимо целлюлозы в состав домика входит и большое количество аминокислот. Кустикообразная форма колоний связана со способом вегетативного деления клеток, когда одна из дочерних клеток, покидая родительский домик, прикрепляется к его отверстию. Вращаясь вокруг своей оси, она формирует собственный домик. Родительский домик могут покинуть и обе дочерние клетки. Клетки содержат 1-2 хлоропласта, на переднем конце клетки видны глазок и 2 сократительные вакуоли. Хризоламинарин расположен в вакуоли на заднем конце клетки. Размножение вегетативное и половое.
Порядок Гиббердиевые – Hibberdiales. Для представителей данного порядка характерен уникальный пигментный состав. Помимо фукоксантина они имеют дополнительный светособирающий каротиноидный пигмент – антераксантин.
Включает роды Гиббердия, Хромофитон, Стилоцерас, Хризопиксис, Платитека и другие.
Род Гиббердия имеет две стадии в жизненном цикле: колониальную пальмеллоидную неподвижную и одноклеточную монадную подвижную. При световой микроскопии у монадной стадии виден только один жгутик. Характерно наличие помимо фукоксантина еще одного дополнительного светособирающего каротиноидного пигмента – антераксантина.
Порядок Гидрурусовые – Hydrurales. Представители данного порядка имеют таллом с пальмеллоидным и псевдопаренхиматозным типами строения. Характерно наличие уникальных зооспор тетраэдрической формы. Глазка нет. Митоз полузакрытый.
Включает роды Гидрурус, Целлониелла, Федерматиум, Хризонебула и другие.
Род Гидрурус (рис. 37, Е) является наиболее дифференцированным среди пальмеллоидных водорослей. Его таллом представляет собой крупные, длиной до 30 см слизистые колонии, имеющие вид коричневых разветвленных шнуров, часто издающих очень неприятный запах. В колониях можно различить главный ствол и боковые разветвления. В клетках, погруженных в общую слизь, содержится один чашевидный хроматофор и несколько сократительных вакуолей. Клетки рыхло расположены по периферии колонии и более плотно в середине. Таллом может расти только за счет деления апикальных клеток. При бесполом размножении в клетках боковых ветвей колонии формируются зооспоры. Образует шаровидные цисты. Встречается в горных ручьях и реках с холодной водой, где прикрепляется к твердым субстратам.
Класс Синуровые водоросли – Sinurophyceae
Название класса происходит от греч. syn – вместе и ura – хвост. Эта группа организмов в 1987 г. выделена из золотистых водорослей. Объединяет монадные одиночные и колониальные организмы, иногда с чередованием в жизненном цикле монадной и пальмеллоидной стадии. Повержность пектиновых оболочек покрыта панцирем из кремниевых чешуек. Митохондрии с трубчатыми кристами, обычно расположены в цитоплазме вокруг хлоропласта. Ядро одно, хлоропластов, окруженных четырьмя мембранами, обычно два. Ламеллы трехтилакоидные, опоясывающая ламелла имеется. Основные пигменты – хлорофиллы а и с, β-каротин и фукоксантин. Стигмы нет. Клетки обычно с 2 неравными жгутиками. Длинный перистый жгутик направлен вперед. Короткий гладкий жгутик, который иногда сильно редуцирован, направлен назад.
Клетки размножаются в основном продольным делением. Колонии, распадаясь, дают начало молодым колониям. У некоторых видов описан половой процесс в форме изогамии. При этом, в отличие от золотистых водорослей, слияние изогамет у синуровых водорослей происходит не на передних, а на их задних концах. Эндогенным путем образуются окремнелые цисты с порой, сходные с таковыми у хризофит.
Преимущественно планктонные формы, пальмеллоидные стадии входят в состав бентоса. Большинство видов синуровых водорослей - фототрофные пресноводные организмы.
Систематика.
Класс Синуровые водоросли включает 7 порядков: Chloramoebales, Synurales, Rhizochloridales, Ochromonadales, Heterogloeales, Parmales и Thallochrysidales.
Порядок Синуровые – Synurales. Монадные формы, пектиновые оболочки клеток обычно покрыты панцирем из кремнеземных чешуек, сцементированных органическим веществом в цельный футляр. Хлоропластов один или два.
Род Синура (рис. 38) - пресноводные монадные формы с двумя неравными жгутиками и двумя пристеночными хлоропластами. Ядро грушевидное, расположено в передней части клетки. Позади ядра имеется одна большая вакуоль, а в задней части клетки – несколько мелких сократительных вакуолей. Клетки покрыты чешуйками, как черепицей Размножение чаще всего вегетативное, для некоторых известен половой процесс. Образует колонии (рис. 39, А). Часто во всех клетках колонии возникают цисты. При массовом развитии Синура может придавать воде неприятный запах.

Рис. 38. Внешний вид клеток Синуры
Порядок Ризохлоридовые – Rhizochloridales. Преимущественно пресноводные организмы с ризоподиальным типом дифференциации таллома, обитающие главным образом в пресных водоемах.
Род Малломонас (рис. 39, Б-Г) – одноклеточная монада с одним хорошо заметным жгутиком и одним раздвоенным хлоропластом.

Рис. 39. Внешний вид синуровых водорослей (по: Г.А. Белякова и др., 2006): А – делящаяся колония Синуры; Б, В – образование статоспоры и Г – внешний вид Малломонаса
Клетка покрыта чешуйками черепитчато и по спирали, некоторые из них несут игловидные шипы (рис. 39, Г). Характерно наличие статоспор (рис. 39, Б, В). Для ряда представителей описан половой процесс - гологамия.
Род Миксохлорис обитает в клетках листьев сфагновых мхов, имеет вид крупного многоядерного плазмодия. Осенью он образует цисты, которые прорастают весной. Из них выходят зооспоры или амёбы, проникающие в пустые клетки листьев и сливающиеся там в плазмодий.
Порядок Хлорамёбовые – Chloramoebales. Включает монадных представителей. Встречаются в соленых и пресных водах.
Род Гетерохлорис – его клетки могут менять форму, образуя псевдоподии. Это явление присуще многим представителям порядка, так же как тенденция к животному способу питания. В клетке имеются несколько хлоропластов, капли масла и хризоламинарин. В передней части клетки имеются сократительные вакуоли, в центре - одно ядро. Размножается вегетативно продольным делением клетки.
Порядок Охромонадовые — Ochromonadales. Объединяет голые формы с двумя неравными жгутиками. Пресноводные и морские формы.
Род Охромонас включает одноклеточные монады с двумя неравными жгутиками. Клетки одеты только плазмалеммой. На переднем конце клетки есть сократительная вакуоль и глазок, на заднем конце — вакуоль с хризоламинарином. Размножается вегетативно, борозда деления начинается у переднего конца клетки между двумя парами жгутиковых оснований. Виды рода распространены в олиготрофных пресные водах, но имеются и морские представители.
Порядок Гетероглеевые — Heterogloeales. Включает водоросли с пальмеллоидным типом дифференциации таллома. Представители порядка встречаются чаще в пресных, чем в соленых водах.
Род Гельминтоглея обитает в солоноватых водах. Эта колония, сидящая на расширенном основании, состоит из ветвящихся слизистых тяжей. В этой слизи беспорядочно располагаются протопласты, каждый из которых окружен собственной слизистой обверткой.
Класс Феотамниевые водоросли – Phaeothamniophyceae
Название класса происходит от типового рода Phaeothamnion (от греч. phaeos – смуглый кустик). Представителей данной группы организмов выделили в отдельный класс из желтозеленых и золотистых водорослей в 1998 г. на основании анализа последовательности рибосомальных генов и особенностей ультраструктуры клетки. Для феотамниевых водорослей характерна уникальная комбинация пигментов: фукоксантина с гетероксантином, отсутствие виолаксантина.
Особенности строения клетки: отсутствие вакуолей с хризоламинарином и отсутствие эндогенных цист с кремнеземными стенками.
Класс включает одноклеточные, колониальные и многоклеточные организмы с коккоидным, пальмеллоидным и нитчатым типом структуры таллома. Жгутиковые стадии с двумя неравными жгутиками. Жгутики латеральные или субапикальные. Стигма встречается у зооспор. Пигменты – хлорофиллы а и с, β-каротин, фукоксантин, диадиноксантин, диатоксантин и гетероксантин. Основным запасным продуктом является парамилон (β-глюкан). Клеточная стенка имеется, во время клеточного деления дочерняя стенка формируется внутри родительской стенки.
У феотамниевых из основных способов размножения встречается лишь вегетативное и бесполое. Бесполое размножение осуществляется автоспорами или зооспорами. Половое размножение неизвестно.
Виды Феотамниевых водорослей обитают исключительно в пресных водах. Систематика.
В настоящее время подтверждена монофилетичность класса феотамниевых водорослей, и среди охрофит он более близок бурым и желтозеленым водорослям, чем золотистым. В настоящее время известно около 30 видов этих водорослей, относимых к одному порядку Феотамниевые – Phaeothamniales.
Род Феотамнион представляет собой прикрепленные ветвящиеся нити до 1 см высоты (рис. 40). Клетки цилиндрические, неправильные, расширенные кверху, по
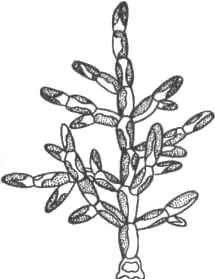
Рис. 40. Внешний вид Феотамниона.
длине в два-три раза превышают ширину. При делении клеток материал родительской стенки расходуется на образование слоистого чехла, окружающего клетки нити. По периферии клетки под плазмалеммой имеются везикулы. Они напоминают физоды – образования, встречающиеся в клетках бурых водорослей. Клетки содержат от одной до нескольких дисковидных пластид оливково-бурого цвета. Зооспоры образуются по 1–2, реже по 4–8 в одной клетке. У зооспор жгутики прикрепляются сбоку. Положение жгутиковых корешков у Феотамниона напоминает таковое у желтозеленых и бурых водорослей. У него также отсутствует ризопласт. В жизненном цикле могут образовываться цисты, но в их стенках отсутствует кремний. Цисты прорастают зооспорами. Поселяется как эпифит на нитчатых водорослях. Обитает в стоячих и медленно текущих пресных водоемах.
Класс Рафидовые водоросли – Raphidophyceae
Название класса происходит от греч. rhaphid – игла. Объединяет одноклеточные двужгутиковые организмы, лишенные клеточной стенки. В основном пресноводные организмы с уплощенными клетками (рис. 41).
Клетки имеют Жгутики неравной длины, направленный вперед жгутик – перистый и длиннее, направленный назад – гладкий и короче. Переходная спираль отсутствует. Хлоропласты мелкие, многочисленные или, реже, по 1-2 в клетке, окружены четырьмя мембранами, из них две мембраны ХЭС. Тилакоиды собраны в стопки по три. Ядро крупное, окружено кольцом диктиосом. Митохондрии с трубчатыми кристами. Имеются сократительные вакуоли. Цитоплазма часто бывает вакуолизирована. Клетки голые, окружены только плазмалеммой. Стигмы (глазка) обычно нет.
Фотосинтетические пигменты – хлорофиллы а и с, β-каротин, вошериаксантин, диноксантин, диадиноксантин, гетероксантин.
Запасные продукты ассимиляции – жиры и масла, реже крахмал и хризоламинарин.
Обитают также в сфагновых болотах и других местообитаниях с кислой и нейтральной реакцией воды. Некоторые виды встречаются в солоноватых и морских водах.
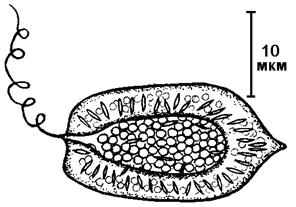
Рис. 41. Внешний вид рафидовой водоросли.
Систематика.
Очень долго эту группу водорослей из-за зеленой окраски, наличия глотки и ряда других особенностей строения рассматривали как класс Хлоромонады среди Евгленовых водорослей. Но данные цитологии, биохимии, физиологии и молекулярной биологии показали их несомненную принадлежность к отделу Гетероконтные (Охрофитовые) водоросли и монофилетичность группы Рафидовых водорослей.
Известно около 25 видов из единственного порядка Хаттонелловые – Chattonellales.
Род Гетеросигма (рис. 42, А) включает морские флагеллаты. Клетки овальной формы со жгутиками, сдвинутыми на боковую сторону, пластиды расположены по периферии клетки. Трихоцист нет. Плавают, совершая вращающие движения вокруг продольной оси тела. Могут вызывать токсичное "цветение воды" в прибрежных водах.

Рис. 42. Рафидовые водоросли (по: R. E. Lee, 1999): А – Гетеросигма; Б – Гониостомум; 1 – хлоропласт; 2 – мукоциста; 3 – сократительная вакуоль; 4 – трихоциста; 5 – ядро
Род Гониостомум (рис. 42, Б) - подвижные дорсовентрально сжатые монады. Спинная сторона выгнутая, брюшная - уплощенная. Борозда на переднем конце ведет в треугольную глотку, из которой выходят два длинных жгутика; их длина сравнима с длиной клетки. Пластиды расположены по периферии клетки. Под плазмалеммой находятся трихоцисты. Обитает в пресных водах с кислотным значением рН.
Класс Евстигмовые водоросли – Eustigmatophyceae
Название класса происходит от греч. eu – «хорошо» и stigma –«метка», «пятно». Класс объединяет голые одноклеточные, реже колониальные, организмы преимущественно коккоидной структуры.
Ядро одно или несколько. Хлоропласт обычно один, окружен 4 мембранами. Жгутик обычно один передний перистый и второе базальное тело или, реже, два неравных жгутика. Стигма (глазок) имеется, расположена за пределами хлоропласта, ее гранулы без мембран.
Рис. 43. Внешний вид евстигмовой водоросли
Хлоропласт обычно единственный, крупный, многолопастной, чашеобразный или париетальный, желто-зеленого цвета. Тилакоиды собраны в ламеллы по три, опоясывающая ламелла отсутствует.
Основные пигменты: хлорофилл а, β-каротин, виолаксантин и вошериаксантин. Пиреноид обычно есть только у вегетативных форм.
Запасные вещества – масла и вещество неустановленной природы, которое откладывается в виде твердого материала вне пластид. Крахмал не образуется.
Обитают в пресных водах, реже встречаются в морях и в почве.
Систематика.
Раньше эту группу организмов относили в Желтозеленым водорослям. В настоящее время Евстигмовые водоросли рассматривают в ранге класса в отделе Гетероконтофиты. Известно около 35 видов, преимущественно пресноводных, относимых к единственному порядку Евстигмовые – Eustigmatales.
Род Евстигматос (рис. 43) – одноклеточные мелкие шаровидные водоросли. Клеточная стенка гладкая, цельная, без орнамента. Желто-зеленый хлоропласт один, лопастной, париетальный, со стебельчатыми многогранными пиреноидами. Большая центральная вакуоль с красным содержимым. Размножается 2 или 4 автоспорами или бутылеобразными зооспорами с одним входящим жгутиком, большой стигмой, расположенной за пределами хлоропласта на переднем конце, и единственным задним хлоропластом без пиреноида. Обычный компонент почвенных флор. Известен из Новой Зеландии, Австрии, Исландии, и из бассейнов Аризоны как "горчичная водоросль".
Класс Желтозеленые водоросли – Xantophyceae
К Желтозеленым относятся водоросли, хлоропласты которых окрашены в светло- или темно-желтый цвет, очень редко зеленый и лишь иногда в голубой. Окраска талломов определяется наличием в хлоропластах клеток следующих пигментов – хлорофиллов а и с, β-каротина и ксантофиллов. Преобладание последних и обусловливает своеобразие окраски желтозеленых водорослей. Кроме того, в клетках в качестве основного продукта ассимиляции накапливаются парамилон, капли масла и лишь у некоторых видов, кроме того, глыбки лейкозина и волютина. Крахмал у желтозеленых водорослей не образуется. Отличительной особенностью желтозеленых является наличие у вегетативных клеток монадной структуры и у зооспор двух неравных жгутиков. В состав клеточной стенки входят целлюлоза, глюкоза и уроновые кислоты. Клеточная стенка часто состоит из двух частей.
Размножение вегетативное, бесполое и половое.
Широко распространены в пресных водах. Редко встречаются в морских, солоноватых водах и в почве.
Ранее класс Желтозеленые водоросли назывался Трибофициевые водоросли по типовому роду Трибонема (от греч. tribon – искусный, хитрый и nema – нить). Известно около 450 видов.
Желтозеленые характеризуются значительным морфологическим разнообразием. Среди многочисленных представителей этого отдела обнаруживаются почти все основные типы структуры тела: амебоидная, монадная, пальмеллоидная, коккоидная, нитчатая, разнонитчатая, пластинчатая и сифональная (рис. 44–46). Слоевище одноклеточное,
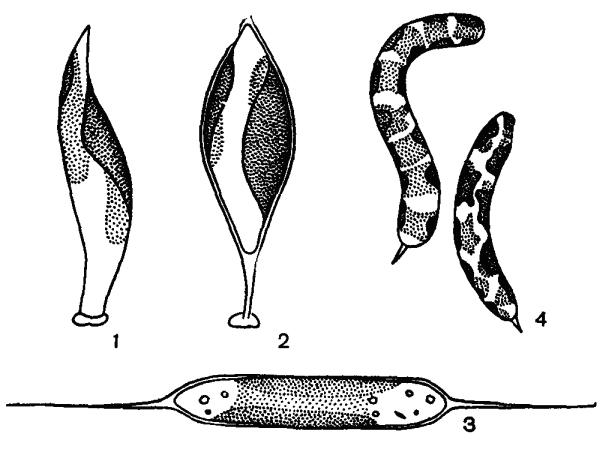
Рис. 44. Внешний вид желтозеленых водорослей: 1, 2 – Харациопсис, 3 – Центритрактус, 4 – Офиоцитиум
колониальное, многоклеточное и неклеточное. Клеточная оболочка плотная, пектиновая и целлюлозная, состоящая из плотно налегающих друг на друга частей или из двух створок. В оболочке откладывается кремнезем или известь. Преимущественно неподвижные формы. Среди одноклеточных видов имеются подвижные формы, лишенные плотной оболочки и снабженные жгутиками, лобоподиями и ризоподиями.
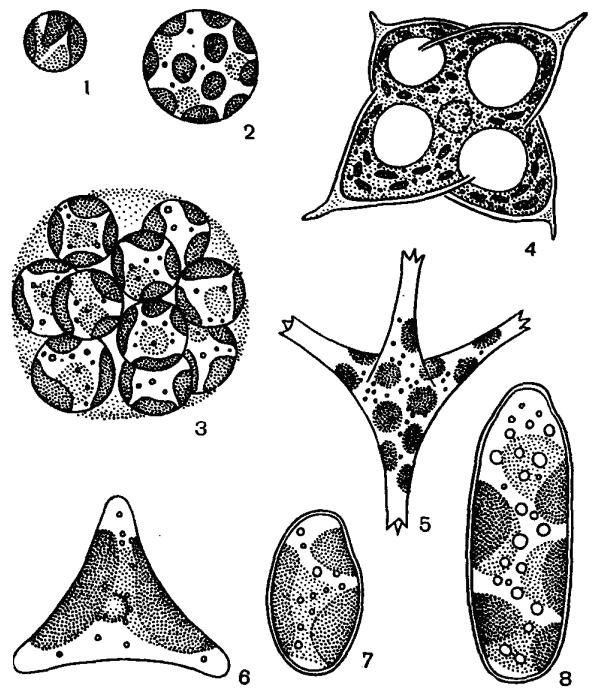
Рис. 45. Внешний вид ксантококковых желтозеленых водорослей: 1–3 – Ботридиопсис, 4 – Тетраэдриелла, 5 – Псевдостаураструм, 6 – Гониохлорис, 7, 8 – Бумиллериопсис
Большинство желтозеленых – неподвижные организмы. У подвижных особей движение может осуществляться с помощью жгутиков либо ризоподий. Клетки различной формы: шаровидные, веретеновидные, эллипсоидные, цилиндрические, тетраэдрические, серповидные, грушевидные, яйцевидные. Талломы размерами от 0.5–1.5 мкм (Хлориделла) до нескольких миллиметров в диаметре (Ботридиопсис) (рис. 45, 1–3) и до десятков сантиметров в длину (Вошерия) (рис. 46, 3).
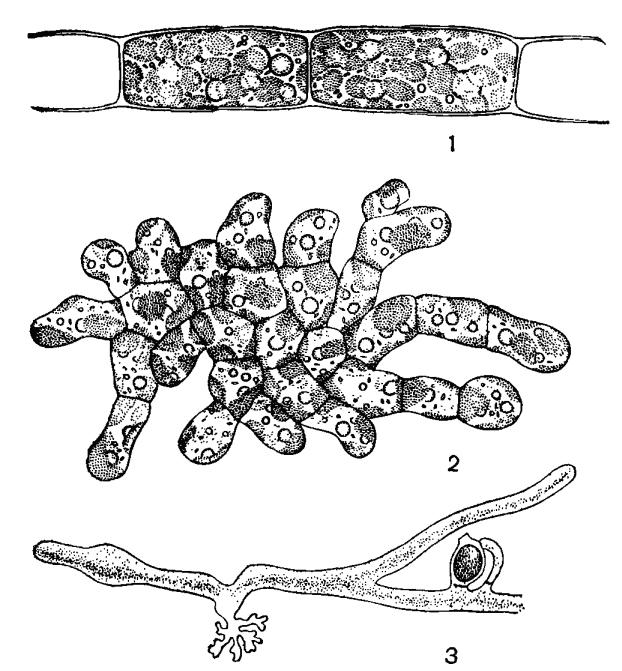
Рис. 46. Внешний вид желтозеленых водорослей: 1 – Трибонема, 2 – Гетеропедия, 3 – Вошерия, часть нити с оогонием и антеридием
Большинство видов желтозеленых являются фототрофами, но встречается и голозойное питание путем заглатывания бактерий и мелких водорослей. Желтозеленые водоросли широко распространены в пресных водах. Они также обычны в почве, реже встречаются в морских и солоноватоводных водоемах. Класс включает аэробионтные, планктонные, донные и перифитонные формы. Эпифиты, эпизоиты, а также внутриклеточные симбионты в клетках простейших.
Независимо от внешней структуры, внутреннее строение клетки желтозеленых водорослей однотипно. В протопласте наблюдается обычно несколько желто-зеленых хлоропластов, имеющих дисковидную, корытовидную, пластинчатую, реже лентовидную, звездчатую или чашевидную форму с цельными или лопастными краями. Окраска связана с отсутствием фукоксантина, отвечающего за золотистый и коричневый цвет у других охрофитовых. Из других пигментов у них имеются β-каротин, вошериаксантин, диатоксантин, диадиноксантин, гетероксантин. У подвижных форм на переднем конце хлоропласта обычно расположен красный глазок, или стигма. У немногих видов имеются пиреноиды полупогруженного типа. Ядро в клетке обычно одно, небольших размеров, но имеются виды и с многоядерными клетками. У некоторых видов в передней части клетки имеется одна или две сократительные (пульсирующие) вакуоли.
У монадных представителей и у подвижных стадий (зооспор и гамет) имеются два неравных жгутика. Исключением являются синзооспоры Вошерии, у которых по поверхности расположены многочисленные пары немного различающихся по длине гладких жгутиков. Короткий жгутик заканчивается акронемой. Жгутики прикрепляются на клетке субапикально. У сперматозоида Вошерии прикрепление латеральное.
У видов с амебоидной, монадной и пальмеллоидной организацией клеточная стенка отсутствует, они покрыты только цитоплазматической мембраной и могут легко менять форму. Иногда «голые» клетки находятся внутри домиков, стенки которых могут быть окрашены в бурый цвет солями марганца и железа. У подавляющего большинства форм имеется клеточная стенка, состоящая из двух частей. В составе клеточной стенки преобладает целлюлоза, а также содержатся полисахариды, состоящие в основном из глюкозы и уроновых кислот. У молодых клеток оболочка тонкая, с возрастом она утолщается. В ней могут откладываться соли железа, соединения которого окрашивают ее в различные оттенки коричневого и красного тонов. Чаще в клеточной стенке присутствует кремнезем, придавая ей твердость и блеск. Она может инкрустироваться также известью и быть различным образом скульптурирована (шипики, ячейки, бородавки, щетинки, зубчики и т.д.) У прикрепленных форм может образовываться вырост оболочки – ножка с прикрепительной подошвой.
У нитчатых форм желтозеленых водорослей с двухстворчатыми оболочками при распаде нитей клеточные оболочки разваливаются на Н-образные фрагменты. Эти фрагменты представляют собой плотно соединенные половинки оболочек двух соседних клеток (рис. 47). При росте нитей Н-образный фрагмент клеточной стенки двух соседних дочерних клеток встраивается между двумя половинками оболочки материнской клетки. В результате каждая из дочерних клеток наполовину покрыта старой оболочкой материнской клетки и наполовину – новообразованной оболочкой.
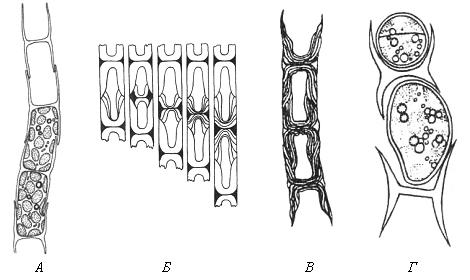
Рис. 47. Схема образования поперечной перегородки между двумя дочерними клетками у нитчатых желтозеленых водорослей (по: А.А. Масюк, 1993): А – фрагмент нити; Б – закладка пояскового кольца и образование поперечной перегородки между двумя клетками; В – слоистость двустворчатых клеточных оболочек; Г – распад оболочки на Н-образные участки
Сократительные вакуоли имеются у подвижных представителей. Обычно их 1-2 на клетку. Аппарат Гольджи своеобразного строения. Диктиосомы мелкие, содержат 3-7 цистерн. Ядро одно, реже их много; у ценотических видов клетки всегда многоядерные.
Размножение. Для большинства видов желтозеленых водорослей характерно вегетативное и бесполое размножение.
Вегетативное размножение осуществляется различными способами: делением клеток пополам, распадом колоний и многоклеточных слоевищ на части. У Вошерии образуются специальные выводковые почки.
При бесполом размножении могут формироваться разнообразные споры: амебоиды, зооспоры, синзооспоры, автоспоры, гемизооспоры, гемиавтоспоры, апланоспоры. Зооспоры «голые» и обычно грушевидной формы.
Половой процесс – изогамия, гетерогамия и оогамия – описан у немногих представителей. У Трибонемы гаметы сходны по размерам, но различаются поведением – это изогамия. У Вошерии наблюдается оогамия: на нитях образуются вместилища женских гамет – оогонии и мужских – антеридии.
В неблагоприятных условиях наблюдается образование цист. Цисты (статоспоры) эндогенные, одноядерные, реже многоядерные. Их стенка часто содержит кремнезем и состоит из двух неравных или, реже, равных частей.
Систематика.
В конце XIX – начале XX вв. различные роды желтозеленых относили к отделу зеленых водорослей, что было связано в первую очередь с окраской и морфологическим сходством талломов. В настоящее время желтозеленые рассматриваются как класс внутри отдела охрофитовых.
Известно около 450 современных видов класса Желтозеленые водоросли, которые группируются в четыре порядка: Ботридиевые, Мишококковые, Трибонемовые и Вошериевые. В основу выделения порядков положены тип дифференциации таллома и особенности жизненного цикла.
Порядок Ботридиевые – Botrydiales. К порядку относятся виды с сифональным типом дифференциации таллома, у которых отсутствует оогамный половой процесс.
Род Ботридиум обитает на почве и имеет вид зеленых пузырей величиной несколько миллиметров, прикрепленных с помощью бесцветных ризоидов. Таллом сифональный, содержит многочисленные ядра и пластиды. Оболочка многослойная, на ней может откладываться известь. Размножение бесполое с помощью двухжгутиковых зооспор, при этом все содержимое пузыря распадается на одноядерные фрагменты. При недостатке влаги размножается с помощью апланоспор или формирует толстостенные цисты. В одних случаях все содержимое пузыря идет на образование одной крупной цисты. В других случаях цисты формируются в ризоидах, куда вначале переходит содержимое пузыря. Цисты прорастают или непосредственно в новый таллом, или образуют зооспоры. Половой процесс – изо- и гетерогамия. Зигота прорастает немедленно, без периода покоя. Обычные и широко распространенные в наземных местообитаниях виды, встречаются по берегам ручьев, прудов или на лишенных растительности почвах.
Порядок Мисхококковые – Mishococcales. Одноклеточные или колониальные представители с коккоидным типом дифференциации таллома.
Род Харациопсис включает одноклеточные прикрепленные формы. При размножении формирует зооспоры, апланоспоры и толстостенные цисты (рис. 44, 1-2).
Род Офиоцитиум (рис. 44, 4) имеет удлиненные цилиндрические клетки, которые могут быть прямыми, согнутыми или спирально закрученными, а на конце могут нести шип. Клеточная стенка состоит из двух неравных частей, из них большая часть участвует в росте клетки, меньшая часть постоянна и имеет форму крышечки. Одноклеточные и колониальные виды, свободноживущие или прикрепленные к субстрату с помощью маленькой ножки. Размножаются зооспорами и апланоспорами, встречаются цисты. Обитают в пресных водах.
Род Мисхококкус образует древовидные прикрепленные колонии. Ветвление дихотомическое и тетрахотомическое. Клетки расположены по 2 или 4 на верхушках слизистых ветвей колонии. Клетки шаровидные, до овальных, с тонкой или толстой клеточной стенкой. Иногда клеточная стенка блестящая и коричневая из-за пропитки ее солями железа. Молодые одноклеточные организмы со слизистым дисковидным основанием, которое служит точкой опоры при прикреплении. После выхода спор протопласт материнской клетки превращается в студень и вытягивается, длина становится в 6 раз больше ширины, и таким образом возникает цилиндрическая ножка. Пустая клеточная стенка материнской клетки всегда становится основанием ножки. Бесполое размножение зооспорами и автоспорами. Автоспоры прикрепляются к верхнему краю слизистой ножки. Последующие клеточные деления повторяют процесс и производят древовидную колонию. Половой процесс – изогамия. Обитают в небольших пресных водоемах как эпифиты нитчатых водорослей. Известны в центральной Европе и Азии.
Порядок Трибонемовые – Tribonematales. Представители имеют нитчатый, разнонитчатый, ложнотканевый и тканевый тип дифференциации таллома. Клеточные стенки либо с Н-образными перекрывающимися частями, либо цельные.
Род Трибонема – неветвящиеся нити (рис. 46, 1). Клетки по форме цилиндрические или бочонкообразные. Клеточная стенка состоит из двух половинок, которые находят краями друг на друга в середине клетки. Оболочки часто слоистые. Фрагменты нитей всегда оканчиваются пустыми половинками Н-образных фрагментов оболочки, имеющих вид вилочки. В клетках несколько желтовато-зеленых пластид, пиреноидов нет. Размножение вегетативное (фрагментацией нитей), бесполое (зооспорами и апланоспорами) и половое (изогамия), причем апланоспоры образуются чаще, чем зооспоры. Могут формировать акинеты. Обитают в пресных водах, где особенно обильно развиваются в холодное время года.
Порядок Вошериевые – Vauсheriales. Все представители имеют сифональный таллом, оогамный половой процесс и синзооспоры.
Род Вошерия (рис. 46, 3) имеет слоевище неклеточного строения; ее таллом достигает в длину нескольких сантиметров, прикрепляется к субстрату с помощью бесцветного ризоида. В нитях отсутствуют перегородки, большая часть таллома занята вакуолью, а по периферии в цитоплазме расположены многочисленные ядра и пластиды. Нити с апикальным ростом и редким боковым ветвлением. Перегородки образуются при повреждении таллома и для отделения органов размножения. Бесполое размножение осуществляется апланоспорами, синзооспорами, акинетами. Синзооспоры формируются по одной в зооспорангии, который отделяется от вегетативных клеток перегородкой на конце нити. Зооспоры многоядерные и многожгутиковые. Половой процесс – оогамия. Зигота покрывается толстой оболочкой и после периода покоя прорастает в новый таллом.
Виды Вошерии широко распространены в пресных, солоноватых и морских водах, а также в наземных местообитаниях. Они встречаются на всех континентах, включая Антарктиду. Формируют травянисто-зеленые или темно-зеленые перепутанные массы – так называемые маты, гладкие, стелющиеся или подушковидные. Водные, полуводные, наземные формы. Обитают в разнообразных местообитаниях: морях, эстуариях, лиманах, соленых маршах, мангровых зарослях, ручьях, каналах, озерах, прудах, на пахотных землях и болотах.
Значение гетероконтных водорослей
Из отдела Гетероконтные водоросли наибольшее значение для природных экосистем и для человека имеют бурые водоросли.
Бурые водоросли – основной источник органического вещества в прибрежной зоне морей. Их биомасса в морях умеренных и приполярных зон может достигать нескольких десятков килограммов на квадратный метр. Заросли бурых водорослей создают условия для питания и размножения множеству прибрежных животных и других водорослей. Чарльз Дарвин, наблюдавший у берегов Южной Америки заросли бурой водоросли Макроцистиса, писал: «Эти огромные подводные леса Южного полушария я могу сравнить только с наземными лесами тропических областей. И все-таки, если бы в какой-нибудь стране уничтожить лес, то не думаю, чтобы при этом погибло хотя бы приблизительно такое количество видов животных, как с уничтожением этой водоросли».
Заросли бурых водорослей служат местом питания, укрытия и размножения многих животных. Образно говоря, бурые водоросли предоставляют другим водным организмам «стол, кров и ясли».
Широко используются бурые водоросли и человеком. Они богаты йодом и другими микроэлементами. Народы Юго-Восточной Азии традиционно используют их в пищу, особенно представителей порядка ламинариевых, из которых готовят множество самых разнообразных блюд. Кормовая мука, приготавливаемая из бурых водорослей, повышает продуктивность скота; при этом в яйцах и молоке увеличивается содержание йода.
Из бурых водорослей получают альгинаты – соли альгиновой кислоты. Альгинаты очень широко применяются в различных областях промышленности. Это нетоксичные соединения, обладающие коллоидными свойствами, поэтому они широко используются в пищевой и фармацевтической промышленности. Альгиновая кислота и ее соли способны к 200–300-кратному поглощению воды, образуя гели, для которых характерна высокая кислотоустойчивость. В пищевой промышленности они используются преимущественно в качестве эмульгаторов, стабилизаторов, желирующих и влагоудерживающих компонентов. Например, сухой порошковый альгинат натрия используют в производстве порошкообразных и брикетированных растворимых продуктов (кофе, чай, сухое молоко, кисели и др.) для их быстрого растворения. Водные растворы альгинатов используют для замораживания мясных и рыбных продуктов. В мире в пищевую промышленность идет до 30% всего объема получаемых альгинатов.
В текстильной и целлюлозно-бумажной промышленности альгинаты используют для загущения красок и усиления прочности их связи с основой. Пропитка тканей некоторыми альгинатами придает им защитные свойства: водонепроницаемость, кислотоустойчивость и увеличивает механическую прочность. Ряд солей альгиновых кислот используют для получения искусственного шелка. Во времена Второй мировой войны в США и Англии из альгиновой кислоты и ее солей производилось большое количество маскировочной ткани и сетей для жилых и промышленных зданий.
Альгинаты применяют в металлургии: в литейном производстве они улучшают качество формовочной земли. Соли альгиновой кислоты применяют в производстве электродов для электросварки, позволяющих получать более высококачественные сварные швы. Альгинаты также используют при производстве пластмасс, синтетических волокон, лакокрасочных покрытий и строительных материалов, устойчивых к атмосферным воздействиям. Их применяют при изготовлении высококачественных смазочных материалов для машин. В радиоэлектронике альгинаты выполняют роль связующего агента при изготовлении высококачественных ферритов.
Наибольшее применение находит водорастворимый альгинат натрия, способный к образованию вязких растворов. Его широко используют для стабилизации разнообразных растворов и суспензий. Добавление небольшого количества альгината натрия в пищевые продукты – консервы, мороженое – повышает их качество. Он также используется для изготовления декоративной косметики, кремов и масок в парфюмерной промышленности.
В фармацевтической промышленности альгинаты используются для покрытия таблеток, пилюль, в качестве компонентных основ для различных мазей и паст, как гели-носители лекарственных препаратов, в производстве растворимых хирургических нитей. В медицине альгинат кальция применяют в качестве кровеостанавливающего средства и как сорбент, выводящий радионуклиды (например, стронций). Ежегодное производство альгинатов в мире превышает 20 тысяч тонн.
Другое важное вещество, получаемое из бурых водорослей, – шестиатомный спирт маннит. Маннит используют как заменитель сахара для больных диабетом. Кроме того, он может быть использован в качестве плазмозаменителя при консервации крови. Его применяют для изготовления таблеток в фармацевтической промышленности. Маннит также используют в производстве синтетических смол, красок, бумаги, взрывчатых веществ, при выделке кож.
Фукоиданы, получаемые из бурых водорослей – эффективные антикоагулянты, даже более активные, чем гепарин. Перспективным считается использование их для получения противоопухолевых препаратов и антивирусных соединений. Ведь даже в самых низких концентрациях фукоиданы могут ингибировать прикрепление вирусов к поверхности клеток. Фукоиданы также обладают способностью образовывать исключительно прочные и вязкие слизи, что находит применение в получении стабильных эмульсий и суспензий.
 Энергетический
кризис, который охватил в последние
годы многие страны мира,
привел к необходимости поиска новых
нетрадиционных источников энергии.
Так, в США с этой целью изучается
возможность разведения гигантской
ламинариевой водоросли Макроцистис
с
последующей переработкой ее в метан.
Подсчитано, что с площади
400 км2,
занятой этой водорослью, можно получить
620 млн. м3
метана.
Энергетический
кризис, который охватил в последние
годы многие страны мира,
привел к необходимости поиска новых
нетрадиционных источников энергии.
Так, в США с этой целью изучается
возможность разведения гигантской
ламинариевой водоросли Макроцистис
с
последующей переработкой ее в метан.
Подсчитано, что с площади
400 км2,
занятой этой водорослью, можно получить
620 млн. м3
метана.
Гетероконтные водоросли из классов Золотистые, Желтозеленые, Синуровые, Рафидофитовые и Евстигмовые водоросли представленные, в основном, микроскопическими организмами, широко распространены в пресных водоемах всех климатических зон земного шара, но чаще встречаются в умеренных широтах. Есть среди золотистых водорослей виды, которые обитают в морях и соленых озерах, а очень немногие – в загрязненных водах. Максимума развития золотистые водоросли достигают в холодное время года: в планктоне они доминируют ранней весной, поздней осенью и зимой. В это время они играют значительную роль как продуценты первичной продукции и служат пищей организмам зоопланктона.
Некоторые золотистые водоросли, например Уроглена и Динобрион, развиваясь в массовом количестве, способны вызывать цветение воды. Они выделяют альдегиды и кетоны, которые могут придавать воде неприятный запах и вкус, a Уроглена – токсичные для рыб жирные кислоты.
Рафидовые водоросли широко представлены в планктоне пресных водоемов преимущественно с кислым рН, особенно в сфагновых болотах, реже в крупных озерах. В пресных водоемах локальные «цветения» может формировать Гониостомум. Рафидовые водоросли встречаются также в опресненных морских бухтах и лужах на берегу моря, а также в открытом море. При массовом развитии в прибрежных морских водах они вызывают токсичное "цветение" воды. Так, у побережья Канады во время "цветения" концентрация клеток рафидовой водоросли Гетеросигмы может достигать 30 млн. на 1 л. Вспышки численности рафидовых водорослей часто приводят к развитию «красных приливов», с которыми связывают гибель рыб. Причиной таких «красных приливов» могут быть виды родов Хаттонелла, Олистодискус, Гетеросигма и Фиброкапса.
Синуровые водоросли при массовом развитии в пресных водоемах могут придавать воде неприятный запах (Синура). Феотамниевые водоросли встречаются в стоячих и медленно текущих пресных водоемах, где поселяются эпифитно на нитчатых водорослях.
Евстигмовые водоросли встречаются только в пресных водоемах или в почве.
Желтозеленые водоросли распространены на всех континентах, они обитают преимущественно в пресных водах и почве, а также в наземных, солоноватоводных и морских местообитаниях. Желтозеленые водоросли населяют чистые и загрязненные воды, с различным значением рН: могут обитать как в кислых, так и в щелочных водах. Они встречаются, главным образом, в чистых пресноводных водоемах, реже в морях и солоноватых водах, предпочитая умеренную температуру, чаще развиваются весной и осенью, хотя имеются виды, встречающиеся на протяжении всех периодов года, в том числе и зимой. Чаще всего их можно найти в скоплениях нитчаток и среди зарослей высших водных растений в прибрежной зоне рек, прудов, озер и водохранилищ.
Подавляющее большинство желтозеленых – свободноживущие формы, но встречаются и внутриклеточные симбионты – зооксантелы в клетках простейших. Интересный внутриклеточный симбиоз образуют хлоропласты морского вида Вошерии с моллюском Элизия. На протяжении девяти месяцев этот моллюск способен в культуре к фотоавтотрофной фиксации углекислого газа. Это самый длительный во времени симбиоз такого типа, когда симбиотическая пластида находится в непосредственном контакте с цитоплазмой животного. В природе личинки моллюска питаются нитями Вошерии. В результате фагоцитоза хлоропласты водоросли попадают в цитоплазму эпителиальных клеток моллюска. В ходе этого процесса оболочка хлоропласта становится трехслойной, теряется одна наружная мембрана ХЭС (хлоропластной эндоплазматической сети). Подобное явление служит хорошим доказательством того, что в ходе эволюции в результате вторичного симбиогенеза за счет потери мембран могли возникать хлоропласты с тремя мембранами.
Желтозеленые, золотистые и другие гетероконтные водоросли являются продуцентами кислорода и органических веществ; они входят в состав пищевых цепей. Гетероконтофиты участвуют в самоочищении загрязненных вод и почв, образовании сапропеля, в процессе накопления органических веществ в почве, влияя на ее плодородие. Они используются как показательные организмы при определении состояния загрязнения вод; желтозеленые водоросли входят в состав комплекса микроорганизмов, применяемого для очистки сточных вод.
Контрольные вопросы
Назовите характерные черты строения бурых водорослей.
Особенности строения талломов бурых водорослей.
Как размножаются бурые водоросли? Что такое моноспоры, тетраспоры и зооспоры, изогамия, гетерогамия и оогамия?
Каковы жизненные циклы у бурых водорослей? Размножение фукусовых и ламинариевых водорослей.
Назовите характерные особенности и типичных представителей порядков бурых водорослей.
В каких местообитаниях встречаются бурые водоросли? Каково значение их в природе.
Хозяйственное значение бурых водорослей.
Назовите характерные черты строения и типичных представителей золотистых водорослей.
Какие пигменты и типы питания известны у золотистых водорослей?
Размножение и экология золотистых водорослей.
Назовите характерные особенности и типичных представителей желтозеленых водорослей.
Какие пигменты и типы питания известны у желтозеленых водорослей?
Как размножаются желтозеленые? Виды полового размножения: изогамия, гетерогамия и оогамия?
Назовите характерные черты строения и типичных представителей синуровых водорослей.
Размножение и экология синуровых водорослей.
Назовите характерные черты строения и типичных представителей феотамниевых водорослей.
Размножение и экология феотамниевых водорослей.
Назовите общие черты строения и типичных представителей рафидофитовых водорослей.
Размножение и экология рафидофитовых водорослей.
Назовите общие черты строения и типичных представителей евстигмовых водорослей.
Размножение и экология евстигмовых водорослей.
Значение гетероконтных водорослей в природных экосистемах.