

5.1. Прокариотические водоросли
5.1.1. Отдел синезеленые водоросли (цианобактерии) – cyanophyta
В названии отдела (от греч. cyanos – синий) отражена характерная особенность этих водорослей – окраска таллома, связанная с относительно высоким содержанием синего пигмента фикоцианина. Цианофиты обычно имеют специфический сине-зеленый цвет. Однако их окраска может сильно варьировать в зависимости от комбинации пигментов – быть почти зеленой, оливковой, желтовато-зеленой, красной и др. В последние годы для синезеленых водорослей все чаще используют другое название – "цианобактерии". Это название лучше отражает две важнейшие характерные черты этих организмов – прокариотическую природу клеток и тесную связь с эубактериями. С другой стороны, традиционное название указывает на такие черты, как способность к оксигенному фотосинтезу и сходство между структурой синезеленых водорослей и структурой хлоропластов эукариот.
Известно около 2 тысяч видов цианофит, широко распространенных в морских и пресных водах и в наземных местообитаниях.
Клетка синезеленых водорослей прокариотная. Она состоит из клеточных покровов (клеточной стенки) и внутреннего содержимого – протопласта, к которому относятся плазмалемма и цитоплазма с различными структурами: фотосинтетический аппарат, ядерный эквивалент, рибосомы, гранулы и др. (рис. 12).
У синезеленых водорослей отсутствуют органоиды, окруженные мембранами: ядро, хлоропласты и т.д., а также немембранные структуры: микротрубочки, центриоли, микрофиламенты.
Наиболее характерными чертами строения клетки синезеленых водорослей являются:
Отсутствие типичных ядер, окруженных ядерными мембранами; ДНК лежит в центре клетки свободно.
Локализация фотосинтетических пигментов в тилакоидах при отсутствии хлоропластов; тилакоиды содержат хлорофилл а.
Маскировка зеленых хлорофиллов красным – фикоэритрином и синими пигментами – фикоцианином и аллофикоцианином.
ДНК расположена в фибриллярно-зернистой нуклеоплазматической области, не окружена мембраной.
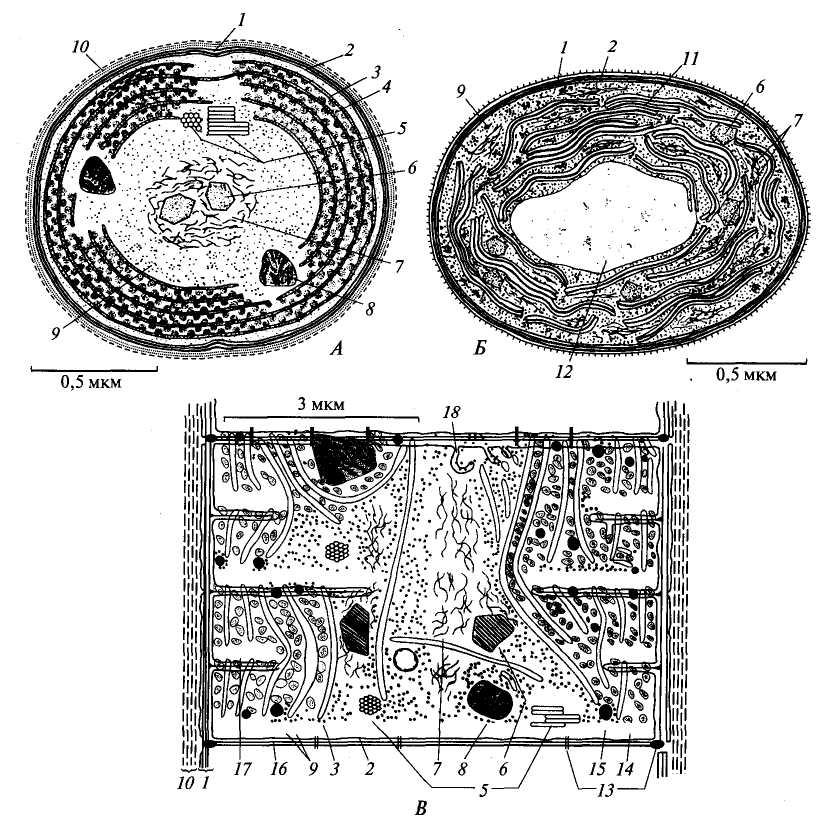
Рис. 12. Строение клетки синезеленых водорослей (по: C.Hoek van den et al., 1995): A – Синехоцистис; Б – Прохлорон; В – Псевдоанабена; 1 – клеточная стенка; 2 – плазмалемма; 3 – тилакоид; 4 – фикобилисома; 5 – газовые везикулы; 6 – карбоксисома; 7 – фибриллы ДНК; 8 – цианофициновая гранула; 9 – рибосомы; 10 – полисахаридный чехол; 11 – стопка тилакоидов; 12 – раздутый тилакоид; 13 – поры; 14 – гранулы цианофицинового крахмала; 15 – липидная капля; 16 – поперечная перегородка; 17 – молодая поперечная перегородка; 18 – впячивание плазмалеммы
Наличие жестких (негибких) слоистых клеточных оболочек.
Образование в большинстве случаев слизистых обверток.
Присутствие разнообразных включений: газовых вакуолей (обеспечение плавучести), цианофициновых гранул (фиксация азота), полифосфатных тел (фиксация фосфора).
Общая характеристика
Для одноклеточных синезеленых водорослей характерен коккоидный тип строения тела. У многоклеточных индивидов встречается нитчатая (трихомальная), реже разнонитчатая (гетеротрихальная) форма строения таллома. Очень редко наблюдается определенная тенденция к пластинчатому или объемному расположению клеток. В нитевидных колониях плазматическая взаимосвязь между клетками отсутствует.
Они могут быть прикрепленными или неприкрепленными к субстрату, неподвижными или способными к скользящему движению. Однако жгутики и реснички никогда не образуются. На движение цианофит различным образом влияет освещение. Во-первых, свет определяет направления движения. Движение по направлению к источнику света называется "положительным фототаксисом", в обратном направлении – "отрицательным фототаксисом". Во-вторых, интенсивность света изменяет скорость движения – "фотокинез". В-третьих, резкое увеличение или уменьшение интенсивности света быстро изменяет направление движения – "фотофобия".
Клетки синезеленых водорослей по форме чаще всего шаровидные, бочонковидные или эллипсоидные, реже вытянутые до цилиндрических и веретеновидных, прямые или согнутые. Иногда клетки грушевидной формы. У прикрепленных одноклеточных индивидов, и иногда и у одноклеточных цианоидов, нередко наблюдается гетерополярность клеток. При этом образуются слизистые ножки и диски, которыми они прикрепляются к субстрату.
Индивиды очень часто образуют разнообразные соединения – колонии индивидов, иногда занимающие большие пространства, и продуцируют значительное количество слизи, нередко заметно влияющей на форму и общий облик колоний.
Особи Cyanophyta обычно микроскопические, но колониальные индивиды у ряда видов могут измеряться сантиметрами.
Основными пигментами синезеленых водорослей являются хлорофилл а, каротиноиды (каротин, ксантофилл) и фикобилипротеиды (аллофикоцианин, фикоцианин, фикоэритрин). Последние встречаются в виде специальных структур – фикобилисом, которые располагаются на поверхности тилакоидов.
Синезеленые водоросли способны к различным видам фотосинтеза: оксигенному и аноксигенному. Оксигенный фотосинтез – это процесс фиксации углекислого газа с использованием воды в качестве донора электронов, сопровождающийся выделением кислорода. Протекает в аэробных условиях. Аноксигенный фотосинтез – процесс фиксации углекислого газа с использованием сероводорода или сульфида в качестве донора электронов, сопровождающийся выделением серы. Происходит в анаэробных условиях. В гипергалинных озерах Израиля, где зимой создаются сильно анаэробные условия, использование комбинации оксигенного и аноксигенного фотосинтеза позволяет водоросли рода Осциллатория доминировать в озере круглогодично. В бескислородных условиях в песках приливо-отливной зоны морей происходит фотосинтез с выделением серы или тиосульфата. Многие цианофиты на свету в анаэробных условиях могут фиксировать углекислый газ, используя водород, однако этот процесс идет с низкой скоростью и быстро прекращается.
У синезеленых водорослей известны несколько типов питания:
Облигатный фотоавтотрофный. Могут расти только на свету на неорганическом источнике углерода.
Факультативный хемогетеротрофный. Способны к гетеротрофному росту в темноте, используя органические вещества, и к фототрофному росту на свету.
Фотогетеротрофный. Используют на свету органические соединения как источник углерода.
Миксотрофный. Используют органические соединения в качестве дополнительного источника углерода. Способны и к автотрофной фиксации углекислого газа.
Продуктом фотосинтеза цианобактерий является цианофициновый крахмал. Он откладывается в небольших гранулах, расположенных между тилакоидами. Цианобактерии способны быстро усваивать и накапливать азот в виде цианофициновых гранул, располагающихся обычно около поперечных перегородок клеток. Фосфаты у синезеленых водорослей запасаются в полифосфатных гранулах, а липиды – в виде капель в цитоплазме по периферии клетки.
Размножения. Все живые клетки синезеленых водорослей способны к делению. Деление клеток многоклеточных и колониальных представителей обычно приводит к росту. Деление клеток возможно в одной, двух, трех или многих плоскостях. У многоклеточных форм при продольном делении в одной плоскости возникают нитчатые формы, в двух плоскостях – пластинчатые, в трех – кубические. При делении одноклеточных особей одновременно осуществляется и размножение. Одноклеточные особи цианофит размножаются равным, реже неравным делением. При этом внутренние слои оболочки клетки врастают вовнутрь клетки. В отдельных случаях наблюдается множественное деление содержимого. Митоз и мейоз отсутствуют. Размножение особей вегетативное, реже бесполое. У ряда представителей цианобактерий образуются покоящиеся споры (акинеты). Типичного полового процесса нет.
Вегетативное размножение у коккоидных форм осуществляется простым делением клетки надвое по всевозможным направлениям в зависимости от случайных воздействий среды. В результате образуются две равные, но не равноценные, части, дающие начало двум новым организмам. Деление клетки надвое происходит в одной или нескольких плоскостях. В последнем случае чаще всего образуются колонии.
Множественное деление клетки происходит в том случае, когда деление клетки и ее ядерной области несогласованно. В результате усиленного деления "ядра" клетка становится многоядерной, затем происходит изоляция участков протоплазмы вокруг "ядер" и образуется множество изолированных эмбриональных клеток. Основными факторами, приводящими к повторному и множественному делению клетки цианобактерий, является избыточное питание, вызывающее ее гипертрофированный рост, а также изменение физико-химических условий существования. Гипертрофированный рост вызывает задержку созревания клетки, и далее повторное либо множественное деление.
Одним из путей вегетативного размножения цианофит является фрагментация (распад) их слоевищ. Причиной фрагментации могут быть механические факторы, отмирание части клеток или нарушение существующих между ними тесных связей. У гормогониевых синезеленых водорослей фрагментация происходит путем распада нити на гормогонии благодаря отмиранию некоторых клеток трихома – некроидов. Каждый гормогоний состоит из 2–3 или большего числа клеток, которые с помощью выделяемой ими слизи выскальзывают из слизистого влагалища и, совершая колебательные движения, перемещаются в воде или по субстрату. Каждый гормогоний может дать начало новой особи. Если группа клеток, похожая на гормогоний, одета толстой оболочкой, ее называют гормоцистой. Она выполняет функции размножения и перенесения неблагоприятных условий. У некоторых видов от слоевища отделяются одноклеточные фрагменты, называемые гонидиями, кокками или планококками. Гонидии сохраняют слизистую оболочку, кокки и планококки лишены выраженных оболочек. Как и гормогонии, они способны к активному движению.
Бесполое размножение осуществляется с помощью особых клеток, не имеющих утолщенных оболочек: "экзоспор" и "эндоспор". Экзоспоры образуются путем неравного деления клетки, когда от материнской клетки отпочковывается меньшая по размерам.
При наступлении неблагоприятных условий (высушивание, холод, дефицит питательных веществ) цианобактерии образуют акинеты. Эти крупные толстостенные покоящиеся споры, заполненные запасными продуктами, служат для переживания этих неблагоприятных условий. Акинеты в течение десятков лет могут сохраняться жизнеспособными, например, в осадках озер при отсутствии кислорода.
Систематика
Все современные формы отдела Суаnорhуtа могут быть сгруппированы в один, два или три класса. Если принять представление о 3 основных путях эволюционного развития синезеленых водорослей от исходных коккоидных одноклеточных форм, то можно согласиться с выделением в пределах Cyanophyta трех классов: Chroococcophyceae – хроококковые водоросли, Chamaesiphonophyceae – хамесифоновые водоросли и Hormogoniophyceae – гормогониевые водоросли.
Класс Гормогониевые – Hormogoniophyceae
(Порядки Осциллаториевые, Ностоковые, Стигонемовые -
Oscillatoriales, Nostocales, Stigonematales)
Для видов характерна трихальная форма строения тела особей, а также способность к образованию гормогониев, т.е. специальных фрагментов нитей, способных к активному произвольному движению и прорастанию в новые особи. Индивиды многоклеточные, "простые" или колониальные (с многоклеточными цианоидами). Нити могут быть разветвленными или неразветвленными, ветвление - настоящим и ложным. При настоящем ветвлении происходит ветвление трихома. При ложном ветвлении ветвятся только влагалища. Трихомы бывают однорядными и многорядными, неразветвленными и разветвленными, гомоцитными и гетероцитными. Гомоцитные трихомы состоят из сходных, не дифференцированных по форме и функциям клеток. Гетероцитные трихомы состоят из клеток, неодинаковых по форме, функциям и локализации. Клетки, которые внешне сходны с клетками гомоцитных трихомов, называют вегетативными; резко отличающиеся от них – особыми. К последним принадлежат гетероцисты и акинеты.
Циклы развития нередко сложные, в процессе которых наблюдается ряд морфологически неодинаковых стадий. Кроме того, гормогониевым водорослям свойственна многовариантность развития.
Род Осциллатория (рис. 13, А). Дифференциации клеток по форме, функциям и локализации нет. Нити неразветвленные, однорядные, гомоцитные. Влагалища отсутствуют или имеются.
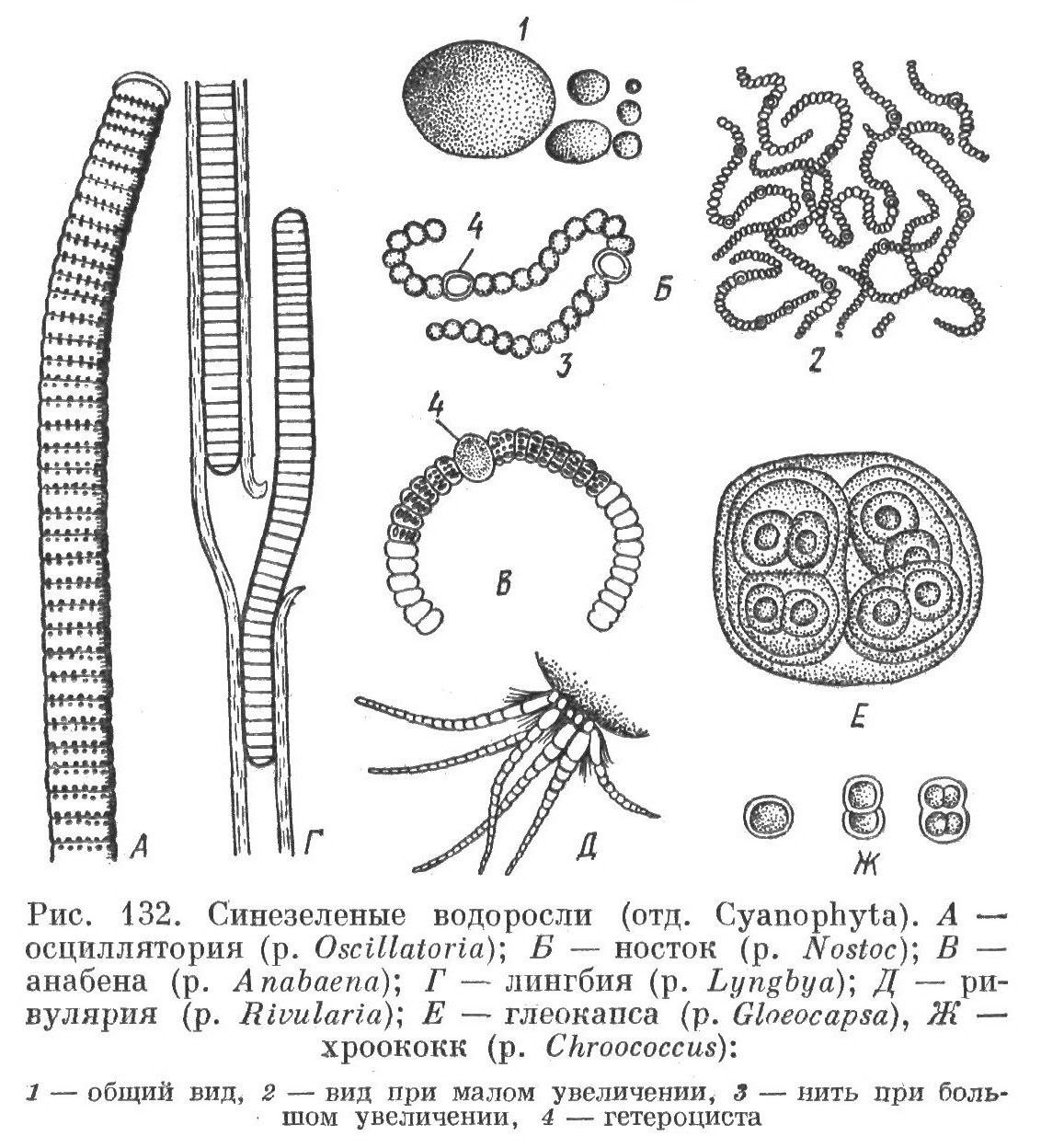
Рис. 13. Морфологическое разнообразие синезеленых водорослей (по: ): А – Осциллатория; Б – Носток; В – Анабена; Г – Лингбия; Д – Ривулария; Е – Глеокапса; Ж – Хроококк: 1 – общий вид, 2 – вид при малом увеличении, 4 – гетероциста
Род Носток (рис. 13, Б). Клетки дифференцированы по форме и функциям. Исключительно колониальные организмы, с хорошо развитой слизью, влияющей на форму колоний. Трихомы гетероцитные, однорядные, неразветвленные, с влагалищами или без них.
Род Ривулария (рис. 13, Д). Слоевище в виде неразветвленных или разветвленных нитей, с влагалищами или без них. Индивиды одиночные или образуют соединения. трихомы гетероцитные, в зрелости асимметричны, суживаются от основы к верхушке, часто оканчиваются волоском, состоящим из вакуолизированных клеток.
Род Стигонема (рис. 14, А). Клетки дифференцированы по форме и функциям. Для видов рода характерно настоящее боковое ветвление. Трихомы гетероцитные, однорядные или многорядные, образуют сплетения, пучки. Нити с влагалищами или, реже, без них. Четкого диморфизма ветвей нет. Главные нити обычно стелющиеся. В старых частях нитей клетки нередко пребывают в глеокапсоидном состоянии: объединены в группы и окружены развитыми слизистыми обвертками.
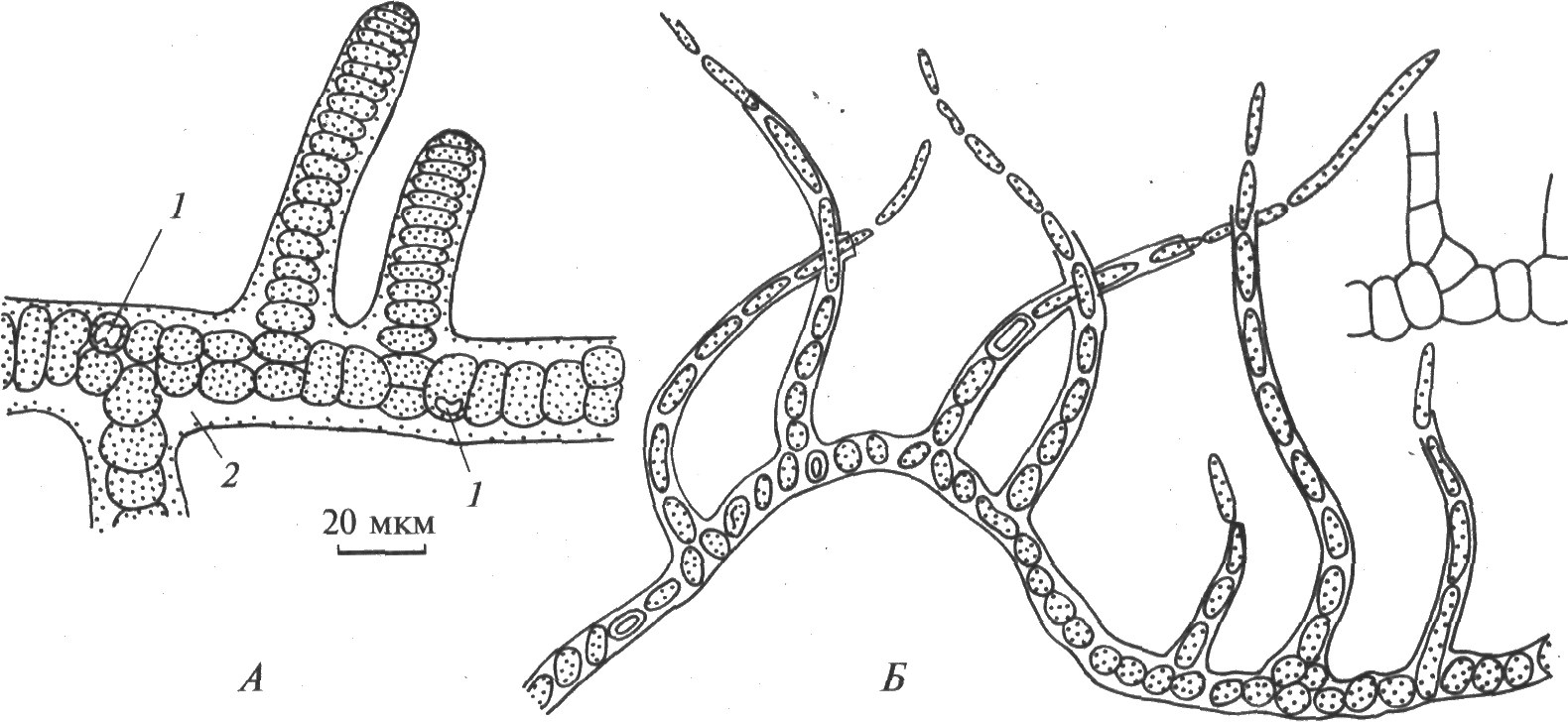
Рис. 14. Стигонемовые синезеленые водоросли (по: R.E. Lee, 1999; М. М. Голлербах и др., 1953): А – Стигонема; Б – Мастигокладус: 1 – гетероциста, 2 – чехол
Род Мастигокладус (рис. 14, Б). Таллом имеет сложное ветвление, гетероцитный. Ветвление истинное и ложное. Клетки основных нитей более или менее шаровидные, клетки ветвей удлиненно-цилиндрические. Влагалища нитей узкие, крепкие или ослизняющиеся. Гетероцисты интеркалярные, споры не известны. Виды рода широко распространены в термальных источниках.
Класс Хроококковые – Chroococcophyceae
Порядок – Chroococcales
Встречаются в виде одноклеточных "простых" индивидов или чаще образуют слизистые колонии. При делении клеток в двух плоскостях возникают однослойные пластинчатые колонии. Деление в трех плоскостях ведет к образованию колоний в виде куба. При делении клеток во многих плоскостях клетки располагаются беспорядочно по всей толщине слизи, а форма колоний разнообразная. Колониальная слизь соединяет простые и сложные цианоиды колоний. Слизь бывает гомогенной или дифференцированной, в виде последовательно вставленных один в другой слизистых пузырей (род Глеокапса) или трубок и тяжей (роды Воронихиния, Гомфосферия). Слизь бывает бесцветная или окрашенная в сине-зеленые, сероватые, оливковые, бурые, красноватые, фиолетовые, черные тона.
Клетки в основном шаровидные или эллипсоидные, реже удлиненные, иногда разнообразно согнутые, цилиндрические или веретеновидные, у некоторых видов яйцевидные, грушевидные или сердцевидные. Хроококковым водорослям свойственно вегетативное размножение. Одноклеточные индивиды делятся надвое в одной, двух, трех или многих плоскостях. Колониальные индивиды размножаются делением колоний и образованием эндогенных колоний. Наиболее часто размножение происходит делением колоний. В пределах этого способа различают фрагментацию колоний, или разрыв их на несколько частей, или перешнуровывание материнской колонии; и почкование колоний, то есть образование на материнской колонии выступов, со временем отделяющихся от нее. Колониальные индивиды также размножаются с помощью обычных вегетативных клеток и спор.
Род Глеокапса образует "простые" или сложные колонии (рис. 13, Е). Клетки шаровидные, эллипсоидные, цилиндрические. Каждая клетка покрыта слизистым чехлом. При делении стенки материнских клеток сохраняются. Колонии округлые или кубические, состоят из последовательно включенных один в другой слизистых пузырей.
Род Микроцистис - колонии сферической или неправильной формы, шаровидные клетки погружены в слизь и могут делиться в любых направлениях (рис. 15). Клетки многих видов содержат газовые вакуоли. Род широко распространен в пресноводном планктоне. Развиваясь в массе, может вызывать цветение воды. Некоторые виды токсичны.
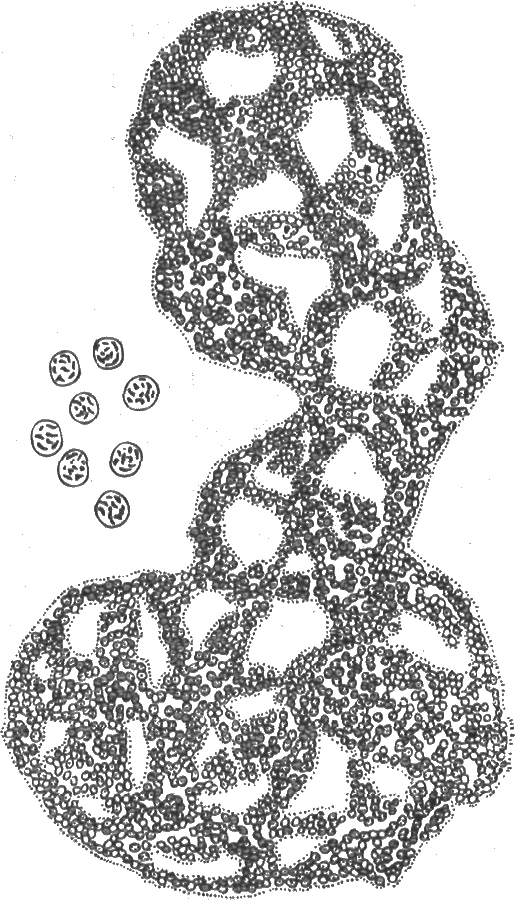
Рис. 15. Хроококковая синезеленая вощоросль Микроцистис (по: М. М. Голлербах и др., 1953)
Класс Хамесифоновые - Chamaesiphonophyceae
(Порядок Плеврокапсовые - Pleurocapsales)
Одноклеточные, часто дифференцированные на основание и верхушку, и колониальные (с одноклеточными цианоидами), обычно прикрепленные к субстрату, индивиды. Характерно образование эндоспор (беоцитов). Клетки разнообразной формы, часто с хорошо выраженными бесцветными или окрашенными в желтоватые или коричневые тона слизистыми обвертками. Деление клеток происходит в одной, двух или трех плоскостях. Клетки в колониях часто очень сжаты и образуют ложную паренхиму, иногда они расположены несколькими слоями. Для многих видов свойственно образование относительно четких рядок клеток, которые напоминают нити. Но плазматическая связь между клетками таких "нитей" отсутствует. "Нити" ползут по субстрату, углубляются в него или поднимаются над ним, при этом нити нередко ветвятся.
Эндоспоры (беоциты) возникают внутри материнской клетки (спорангия), сходной с обычными клетками или отличающейся от них по размеру и форме. Освобождаются беоциты путем разрыва, ослизнения оболочки спорангия или путем сбрасывания части стенки спорангия как крышечки на их образование идет все содержимое спорангия или только его часть.
Род Дермокарпа. Индивиды одноклеточные, дифференцированные на основание и верхушку, прикрепленные к субстрату. Обычно живут одиночно, небольшими группами. Размножаются обычно беоцитами.
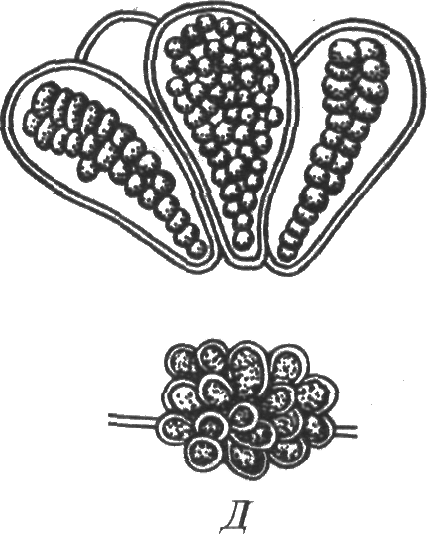
Рис. 16. Хамесифоновая синезеленая водоросль Дермокарпа (по: М. М. Голлербах и др., 1953)
Экология и значение
Синезеленые водоросли распространены повсюду. Их можно найти как в горячих источниках и артезианских колодцах, так и на поверхности снега и влажных скал, на поверхности и в толще почв, в симбиозе с другими организмами: простейшими, грибами, морскими губками, эхиуридами, мхами, папоротниками, голосеменными растениями. Виды синезеленых обычны в планктоне и бентосе стоячих и медленно текучих пресных вод, в солоноватых и соленых водоемах. Они – важные компоненты морского фитопланктона. Синезеленые водоросли играют ключевую роль в океанических экосистемах, где большая часть суммарной фотосинтетической продукции приходится на пикопланктон. Пикопланктон состоит в основном из одноклеточных коккоидных цианофит. Подсчитано, что 20% фотосинтетической продукции океанов дают планктонные синезеленые водоросли. В бентосе встречаются эпифитные, эпилитные и эндолитные формы. Цианобактерии обычно обладают специальными органами прикрепления в виде подошвы, стопы, слизистых тяжей. Обильны также виды синезеленых водорослей, прикрепляющиеся к подводным предметам с помощью слизи.
Цианобактерии являются типичными обитателями горячих вод. Они вегетируют при температуре 35–52°С, а в отдельных случаях до 84°С и выше, нередко при повышенном содержании минеральных солей или органических веществ (сильно загрязненные горячие сточные воды заводов, фабрик, электростанций или атомных станций).
Дно гипергалинных водоемов иногда сплошь покрыто синезелеными водорослями, среди которых преобладают виды родов Формидиум, Осциллатория, Спирулина и др. Синезеленые водоросли обитают на коре деревьев (виды родов Синехококкус, Афанотеке, Носток). Они нередко эпифитируют на мхах, где, например, можно наблюдать черновато-синие пучки видов рода Шизотрикс.
Представители Cyanophyta наиболее обычны среди водорослей, обитающих на поверхности обнаженных скал. Цианофиты и сопутствующие им бактерии образуют "горный загар" (наскальные пленки и корочки) на кристаллических породах различных горных массивов. Особенно обильны разрастания водорослей на поверхности влажных скал. Они образуют пленки и наросты различного цвета. Как правило, здесь обитают виды, снабженные толстыми слизистыми обвертками. Разрастания бывают различного цвета: ярко-зеленые, золотистые, бурые, охристые, лиловые или темно-сине-зеленые, коричневые, почти черные в зависимости от образующих их видов. Особенно характерны для орошаемых скал виды родов Глеокапса, Глеотеке, Хамесифон, Калотрикс, Толипотрикс, Сцитонема.
Представители Cyanophyta составляют подавляющее большинство почвенных водорослей. Они обитают в глубоких и в поверхностных слоях почвы, устойчивы против ультрафиолетового и радиоактивного излучения. В почвах степной зоны Носток обыкновенный образует на поверхности мощные пленки темно-зеленого или, в сухой сезон, корочки грифельно-черного цвета. Массовое развитие микроводорослей вызывает позеленение склонов оврагов, обочин дорог, пахотных почв.
Синезелёные водоросли являются компонентами слоевища многих лишайников, сожительствуют с высшими растениями, например, с водным папоротником Азолла и другими. Как симбионты они защищают своего партнера от высокой интенсивности света, снабжают его органическими веществами, обеспечивают соединениями азота. В то же время от хозяина они получают защиту от неблагоприятных внешних факторов, а также необходимые для роста органические вещества. Лишь немногие симбиотические ассоциации цианофит с различными организмами являются облигатными. Большинство цианофит способны расти самостоятельно, хотя хуже, чем в симбиозе. Они формируют два типа объединений с другими организмами – внеклеточные: с грибами и внутриклеточные: с губками, диатомовыми водорослями и др.
Синезелёные водоросли принадлежат к числу древнейших организмов, их ископаемые остатки и продукты жизнедеятельности обнаружены в породах, образовавшихся 3–3,5 млрд. лет назад, в архейскую эру. Полагают, что первые на Земле (докембрийские) экосистемы состояли только из прокариотических организмов, в том числе цианобактерий. Интенсивное развитие цианофит имело громадное значение для развития жизни на Земле, и не только из-за накопления ими органического вещества, но и в связи с обогащением первичной атмосферы кислородом. Существенную роль cинезелёные водоросли сыграли и в создании известковых пород.
Азотфиксация. Атмосфера Земли на 78% состоит из азота, но способность к его фиксации обнаружена только у прокариот, а среди водорослей исключительно у цианофит. Синезеленые водоросли – уникальные организмы, которые способны к фиксации как углекислого газа, так и атмосферного азота. При фиксации азота выделяется аммиак и водород. Этот процесс обычно происходит в специальных толстостенных клетках со слизистыми чехлами – гетероцистах. Внутри гетероцист создаются условия с малым содержанием кислорода. Фиксация азота днем идет быстрее, чем ночью, так как во время фотосинтеза образуется необходимая для этого процесса АТФ – аденозинтрифосфорная кислота. Путем фиксирования атмосферного азота синезеленые водоросли получают азот, необходимый для синтеза своих белков и дальнейшего роста. Другие водоросли целиком зависят от нитратов и аммония, растворенных в воде.
Биологическая фиксация атмосферного азота является одним из важных факторов повышения почвенного плодородия. Ведущая роль в этом процессе принадлежит цианофитам, которые не требуют для усвоения молекулярного азота готового органического вещества, а сами приносят его в почву. Например, для почв умеренной зоны годовую продукцию азотфиксирующих синезеленых водорослей оценивают в 20-577 кг/га (в сухой массе). К самостоятельной азотфиксации в размерах, имеющих реальное значение для плодородия почвы, способны только гетероцистные формы цианофит (виды родов Носток, Анабена, Калотрикс, Толипотрикс и Цилиндроспермум).
Некоторые представители синезеленых водорослей являются съедобными (Носток, Спирулина). В специальных биологических прудах сообщества синезеленых водорослей и бактерий используют для разложения и детоксикации гербицидов. Некоторые цианобактерии разлагают фенилкарбаматные гербициды на анилин и хлорпроизводные. Сточные воды, очищенные с помощью наиболее совершенных методов, все же остаются токсичными по отношению к гидробионтам. Только альгобактериальные сообщества, которые используются для доочистки сточных вод, позволяют получить воду, соответствующую ГОСТу "Вода питьевая".
"Цветение" воды. Под "цветением" воды понимают интенсивное развитие водорослей в толще воды, в результате чего она приобретает различную окраску, в зависимости от окраски и численности организмов, вызывающих "цветение". Массовому развитию водорослей вплоть до "цветения" воды способствует увеличение евтрофирования водоемов, которое происходит как под влиянием природных факторов (за тысячи и десятки тысяч лет), так и в значительно большей степени под влиянием антропогенных факторов (за годы, десятки лет). "Цветение" воды наблюдается как в континентальных водоемах (пресных, солоноватых и соленых), так и в морях и океанах (в основном, в прибрежных районах). Красное море получило свое название из-за обильного развития в нем синезеленой водоросли Oscillatoria erythraea. Лужеобразные пресные водоемы Средней Европы нередко окрашиваются в красный цвет Haematococcus pluvialis. Из пресноводных водоемов в первую очередь подвержены "цветению" большие равнинные реки и их водохранилища, а также пруды различного назначения, озера, водоемы-охладители.
Умеренная вегетация цианофит положительно влияет на экосистему водоема. При значительном увеличении биомассы водорослей (до 500 г/м3 и выше), начинает проявляться биологическое загрязнение, вследствие чего значительно ухудшается качество воды. В частности, изменяется ее цветность, рН, вязкость, снижается прозрачность, меняется спектральный состав проникающей в водную толщу солнечной радиации в результате рассеивания и поглощения водорослями световых лучей. В воде появляются токсические соединения и большое количество органических веществ, служащих питательной средой для бактерий, в том числе и патогенных. Вода обычно приобретает затхлый неприятный запах. Возникает гипоксия, или дефицит растворенного кислорода; он расходуется на дыхание водорослей и разложение отмершей органики. Гипоксия приводит к летним заморам гидробионтов и замедляет процессы самоочищения и минерализации органики.
Среди цианофит есть патогенные виды (около 30), вызывающие заболевания и гибель рифовых кораллов, при "цветении" воды могут отмечаться заболевания домашних животных и человека, массовая гибель гидробионтов, водоплавающей птицы и домашних животных особенно в жаркие летние месяцы. Отравления людей встречаются значительно реже. Наибольшему риску подвергаются дети и люди с заболеваниями печени и почек. По способу действия токсины цианобактерий разделяют на 4 группы: гепатотоксины, нейротоксины, цитотоксины и дерматотоксины. Они вызывают пищевые интоксикации, аллергии, коньюктивит, поражения центральной нервной системы и т.д. По своему действию цианотоксины в несколько раз превосходят такие яды, как кураре и ботулин. Профилактика чистоты водоемов заключается в недопущении скопления водорослей вблизи водозаборов и мест отдыха или водопоя домашних животных.
"Солнечные реакторы" и водоросли. В последнее время перед человечеством остро встала проблема рационального использования природных энергоресурсов и поиска нетрадиционных источников энергии. К таким источникам относят энергию Солнца, консервируемую в биомассе растений (биоконсервация солнечной энергии). В отличие от атомной энергии, этот источник энергии абсолютно безопасен; использование его не вызывает нарушения экологического равновесия, не ведет к радиоактивному или тепловому загрязнению среды.
Наиболее перспективным считаютиспользование синезеленый водорослей для получения биотоплива путем метанизации биомассы водорослей, выращенных на сточных водах. Установки для получения метана из водорослей созданы в США и Японии. Их продуктивность составляет соответственно 50 и 80 т/га (в сухой массе) в год, а 50-60 т сухой биомассы водорослей может дать 74 тыс. кВт/ч электроэнергии.
Контрольные вопросы
Назовите характерные черты строения клетки цианобактерий.
Какие пигменты и типы питания известны у цианофит?
Как размножаются синезеленые водоросли? Что такое гормогонии, экзоспоры, акинеты?
С какими группами организмов наиболее сходны и когда возникли синезеленые водоросли?
Назовите характерные особенности и типичных представителей синезеленых водорослей класса Хроококковые.
Назовите характерные особенности и типичных представителей синезеленых водорослей класса Гормогониевые.
Назовите характерные особенности и типичных представителей синезеленых водорослей класса Хамесифоновые.
В каких местообитаниях встречаются синезеленые водоросли? Их значение в природе.
Роль цианофит в биологической фиксации атмосферного азота.
Хозяйственное значение цианофит. Оценка качества воды.
Что такое "цветение воды" и цианотоксины?
Синезеленые водоросли как нетрадиционные источники энергии.