

Lektsii_po_biofizike_Baskakov_Kapilevich
.pdf10.Скорость проведения импульса по мякотному нервному волокну зависит от:
A.Диаметра волокна
B.Корня квадратного из диаметра волокна
C.Квадрата диаметра волокна
11.При проведении импульса через зону расширения волокна задержка проведения обусловлена:
A.Уменьшением входного сопротивления
B.Уменьшением удельного сопротивления
C.Увеличением емкости мембраны
12.При проведении импульса через зону расширения волокна задержка проведения обусловлена:
A.Уменьшением первой фазы локального тока
B.Увеличением второй фазы локального тока
C.Уменьшением третьей фазы локального тока
13.При приходе в точку 7-кратного суммарного расширения 2 волокон 2 импульсов с интервалом 1 мсек:
A.Будут проведены оба импульса
B.Будет проведен 1 импульс
C.Проведение будет заблокировано
14.При проведении нервного импульса из мякотной части волокна в безмякотную терминаль задержка проведения обусловлена:
A.Увеличением сопротивления мембраны
B.Увеличением площади мембраны
C.Затратой энергии на выброс медиатора
15.Функциональные неоднородности обусловлены:
A.Изменением удельного сопротивления мембраны
B.Изменением удельной емкости мембраны
C.Изменением свойств белков внешней поверхности мембраны
16.Парабиоз Введенского проявляется:
A.Снижением частоты импульсов
B.Периодическим выпадением импульсов
C.Непериодическим выпадением импульсов
17.Основная ошибка модели парабиоза Беркенблита заключается в следующем:
A.Она не учитывает кинетику инактивации натриевого тока
B.Она не учитывает явления абсолютной рефрактерности
C.Она не учитывает влияние на мембрану импульса, пришедшего в фазу рефрактерности
18.Константы, входящие в кабельное уравнение, зависят от:
A.Амплитуды потенциала действия
B.Ионной проницаемости мембраны
C.Пассивных электрических свойств мембраны
19.Воротный ток – это:
A.Ток, протекающий через канал при закрытых воротах
B.Ток, протекающий через канал в процессе открывания ворот
141
C.Ток смещения, регистрируемый в толще мембраны во время открывания ворот
20.Метод фиксации потенциала используется для:
A.Измерения ионных токов
B.Измерения сопротивления мембраны
C.Измерения мембранного потенциала
Правильные варианты ответов: |
|
|
|
|
|
|
|
1. |
B, C |
6. |
A |
12. |
B |
18. |
C |
2. |
A, C, |
7. |
B |
13. |
B |
19. |
C |
|
D |
8. |
B |
14. |
B |
20. |
A |
3. |
B |
9. |
B |
15. |
A, B |
|
|
4. |
A |
10. |
A |
16. |
B |
|
|
5. |
A |
11. |
A |
17. |
C |
|
|
142

X. БИОФИЗИКА СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
Общие положения
Различают несколько видов синаптической передачи (синапсов):
1.Электрические синапсы.
2.Химический синапс.
3.Смешанные синапсы.
Их функция обеспечить (возбуждающие синапсы) или предотвратить (тормозные синапсы) передачу нервного импульса с одного электровозбудимого объекта (нейрон – нейрон, нейрон – мышца) на другой.
Критерием дифференцировки синапсов являются их отличия касательно электрофизиологических свойств, морфологических характеристик, фармакологической регуляции.
А. Электрофизиологические свойства:
1. Наличие электрической связи между клетками (рис. 48).
Для электрического синапса (ЭС) она обеспечивает зависимость тока входящего направления через мембрану пресинаптической клетки от:
Э.С. |
Vпостсинапт. |
и равна 0,1-0,5 (определяется RC- -мембран); |
Vпресинапт. |
Для химического синапса (ХС) 0.
2. Величина синаптической задержки (интервал между началом пресинаптического ПД и постсинаптической деполяризацией):
Для ХС он равен 0,2 и выше (до 0,8-100) мс. Для ЭС = 0,1 мс (зависит от RC- -мембран).
3. Зависимость постсинаптического потенциала (ПСП) от исходного уровня мембранного потенциала (Vм):
Для ЭС – нет зависимости, так как ПСП определяется Vпресинапт. и коэффициентом передачи.
Для ХС – есть, и величина тока Ii через постсинаптическую мембрану равна:
Ii gi (Vi Vm)
, где
Vi и Vm – уровни ПСП и исходного мембранного потенциала.
Б. Морфологические характеристики:
ЭС отличает малая ширина синаптической щели (100 нм), наличие специализированных структур щелевых контактов. В их состав входят особые каналы, проницаемые для низкомолекулярных веществ (1–2 кД). Щелевые контакты получили название высокопроницаемых контактов (ВПК). Они
143
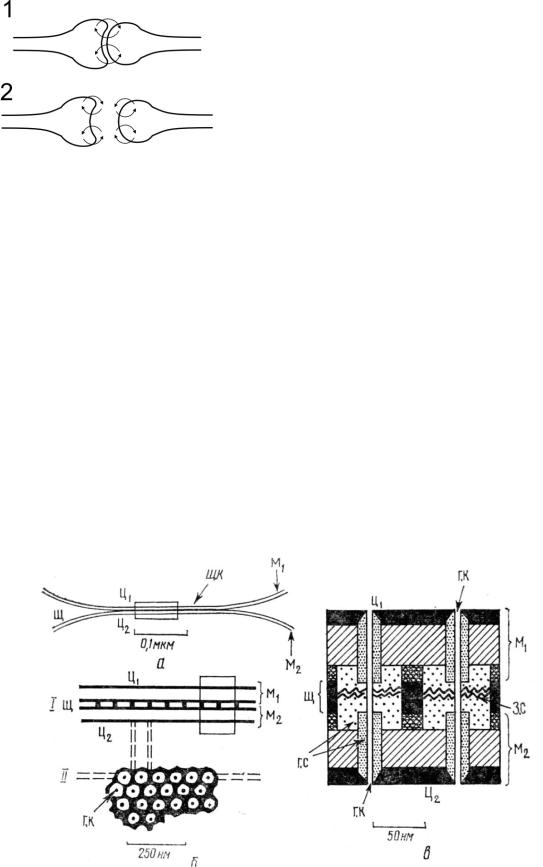
чувствительны к механическому разобщению из-за удаления ионов кальция из раствора, алкалоза, действия гипертонического раствора.
Рис. 48. Токи в электрическом (1) и химическом (2) синапсе.
Химический синапс представляет особую структуру, в состав которой входят пре- и постсинаптическая мембраны (толщина мембран 4–5 нм) и синаптическая щель (100–150 мкм). Пример – двигательная пластинка нервномышечной синаптической передачи. Зона активного контакта концевой пластинки -1,5 мкм. Аксоплазма имеет синаптические пузырьки диаметром
50 нм (рис. 38).
В. Фармакологические критерии:
ЭС – не чувствительны к замене ионов Са2+ на Mg2+, Со2+.
ХС – чувствительны к действию медиаторов и их метаболизму, уровню ионов кальция.
Электрические синапсы
Каждый из видов синаптической передачи имеет преимущества и недостатки. Например, ЭС отличает от ХС то, что они:
1)малозависимы от метаболизма (ферментов);
2)нет истощения функций;
3)проводят высокочастотные серии импульсов.
Рис. 49. Щелевой контакт.
144

Электрические синапсы подразделяют на:
1.Невыпрямляющие синапсы. Когда ток через постсинаптическую мембрану вызывает линейные сдвиги Vмембр. независимо от направления движения импульса.
2.Выпрямляющие. Когда Vpre Vpost и импульс пользуется преимуществом проведения в одном из направлений.
Типичным представителем электрических синапсов является щелевой контакт (рис. 49).
Химический синапс
Условиями функционирования ХС является следующее:
1)медиатор выделяется при возбуждении;
2)должен прийти стимул – эффект возбуждения;
3)блокаторы синаптической передачи должны быть активны;
4)должно быть метаболическое обеспечение медиатора.
Электрические явления в ХС:
1.В состоянии покоя регистрируется миниатюрный потенциал концевой пластинки (МПКП) – электрический эквивалент выделения в синаптическую щель спонтанного выделения 1 кванта медиатора. МКПК = 0,4–0,6 мВ. Зона действия несколько каналов.
2.При стимуляция нейрона 30–50 Гц регистрируется ответ -50мв –
|
потенциал |
концевой |
пластинки |
|
|
(ПКП). |
Он |
соответствует |
|
|
критическому |
|
уровню |
|
|
деполяризации |
|
скелетного |
|
|
мышечного волокна. |
|
|
|
|
Квантово-везикулярная теория |
|||
|
(Бернард Кац и Рикарло Миледи, |
|||
|
Нобелевская премия 2003 года). |
|||
|
Пусть m – квантовый состав |
|||
|
ПКП; n – величина одного кванта; р |
|||
|
– вероятность выделения кванта (m |
|||
|
= pn); где m – целое случайное |
|||
|
число и различие между ответами |
|||
|
кратно n. |
|
|
|
|
Для параметров n |
400 и р |
||
Рис.50. Строение химического синапса. |
0,25 эта теория верна. |
|
|
|
Взаимосвязь пресинаптического и постминаптического потенциалов характеризуется кривой Ленгмюра (рис. 51).
145

Рис. 51. Зависимость пресинаптического потенциала от постсинаптического потенциала концевой пластинки.
Как уже ранее утверждалось, основным электрофизиологическим отличием ХС от ЭС является то, что амплитуда ПКП зависит от Vm скелетной мышцы. Причины такой зависимости в следующем.
1.Исходно амплитуда ПКП всегда на 10-20 мВ ниже ПД мышц. Причиной является постоянный шунт сопротивления мембраны КП, развивающийся под действием на нее нейромедиатора ацетилхолина (АХ). В результате высвобождения АХ в синаптическую щель снижается сопротивление постсинаптической мембраны в 1000 раз.
2.Мембраны КП и мышечного волокна, по сути, одна электрофизиологическая структура (Рис. 52).
Рис. 52. Эквивалентная электрическая схема синапса.
Если ключ К разомкнут, то регистрируется ПП мышцы (-80 мВ). Замыкание К увеличивает проницаемость постсинаптической мембраны. Если е Е, то замыкание ключа вызывает ток в цепи е Е (е -15-0 мВ). Ток бежит по первому пути и начинается деполяризация постсинаптической мембраны до уровня:
V |
g |
е Е |
|
g G |
|||
|
, где |
g и G – проницаемости мембран КП и мышечного волокна, соответственно. Естественно, чем ниже Е, тем выше V (отличие химического синапса).
146

В итоге:
1. При е = Е, ПКП = 0, уровень потенциала реверсии.
2. Если е Е, ПКП меняет свой знак и наступает гиперполяризация постсинаптической мембраны – торможение передачи возбуждения на мышцу.
3. Так как присутствие См шунтирует V и Vm не будет совпадать с развитием постсинаптического мембранного потенциала (ПКП), спад ПКП относительно спада (тока ТКП) концевой пластинки запаздывает и ТКП является характеристикой действия медиатора.
4. Е = е при потенциале реверсии ( реверсии при Ер -15мВ). При этом уровне ПКП тока после действия медиатора не будет.
5. Величина потенциала реверсии рассчитывается по формуле:
реверсии ЕрWa ЕрК
2 , где
ЕрNa и ЕрК – равновесные потенциалы для ионов натрия и калия, соответственно.
6.Нейромедиатор (АХ) одновременно увеличивает проницаемость для Na+
иК+:
gNa 1,29=const на всем протяжении мембраны в сфере действия
gK
нейромедиатора.
Уравнение Т.Takeuchi. Если величина потенциала постсинаптической мембраны зависит только от количества медиатора, то потенциал реверсии будет равен:
Е р RT F
ln
K ext |
|
|
Δg Na |
||
K int |
|
|
Δg K |
||
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
Na ext |
|
|
1 |
|
|
|
|||
lg |
|
|
|
|
||||||
Na |
int |
|
|
1 |
g |
Na |
|
|
||
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
g K |
|||||
|
|
|
|
|
|
|
|
, где |
||
обозначения соответствующих величин концентраций, проводимости и констант объяснялись выше.
Основные положения о судьбе медиатора в химическом синапсе (Шеррингтон Ч., 1897)
1.В холинергическом синапсе производится и утилизируется при действии одного импульса 2-200х103 молекул ацетилхолина (АХ). Возникает вопрос: «Откуда они берутся?».
2.Тем более, что амплитуда МПКП определяется размерами кванта медиатора, объёмом диффузионного пространства, плотностью рецепторов в постсинаптической мембране, активность фермента расщепляющего АХ – ацетилхолинэстеразой (АХЭ), входным сопротивлением мембраны КП.
147

|
|
3. Оказывается, что до взаимодействия с |
|||||
|
КП медиатор претерпевает достаточно сложные |
||||||
|
изменения, протекающие в несколько стадий: |
|
|||||
|
|
a) |
синтез |
и |
загрузку в |
везикулы |
|
|
(пертоикарион, тело нейрона); |
|
|
||||
|
|
б) |
аксональный транспорт; |
|
|
||
|
|
в) |
слияние |
везикулы с пресинаптической |
|||
|
мембраной и высвобождение медиатора в |
||||||
|
синаптическую щель; |
|
|
|
|||
|
|
г) |
диффузия через синаптическую щель; |
|
|||
|
|
д) |
узнавание, |
взаимодействие |
с |
||
|
рецепторами постсинаптической |
мембраны, |
|||||
|
изменение проницаемости ионов КП (рис. 41); |
|
|||||
|
|
е) |
расщепление АХ ацетилхолинэстеразой; |
|
|||
|
|
ж) |
ресинтез ацетилхолина, который связан |
||||
|
с активностью АцетилКоА, гликолизом холина из |
||||||
|
фосфотидилхолина: |
|
|
|
|||
Рис.53. Модель Карлин (ионный |
АцетилКоА + холин =АХ (тело нейрона); |
|
|||||
канал холинергического |
|
з) загрузка синаптических пузырьков АХ |
|||||
синапса). |
|
с |
помощью активного транспорта, когда |
||||
концентрация АХ в них становится больше 5мМ; |
|
|
|
||||
и) |
диффузия АХ от тела нейрона по аксону к месту высвобождения |
||||||
происходит с помощью олигосахаридного матрикса цитоскелета; к) высвобождение нейромедиатора, связанное с ионами кальция и КМ из
везикулы прямо в синаптическую щель.
148

XI. БИОФИЗИКА СОКРАТИТЕЛЬНЫХ СИСТЕМ
Введение
Подвижность и перемещение живых структур имеет много общих черт. В их основе лежат сходные по своей структуре и функции сократительные белки актина и миозина, взаимодействие которых, в принципе, осуществляют необходимые перемещения в пространстве. Существуют специализированные структуры, выполняющие сократительные функции, – мышечная ткань, хотя сократительные белки (актин) являются обязательным компонентом цитоскелета любой клетки.
Виды мышечного сокращения: Изометрическое – при неизменных размерах. Изотоническое – при неизменной силе.
Ауксотоническое – при изменении длины и силы.
Мышечная ткань подразделяется на:
1.Поперечно-полосатую (скелетная и сердечная) мускулатуру.
2.Гладкие мышцы.
Их отличия касаются структуры, функции и эмбриогенеза.
Скелетные мышцы
Структурно-функциональной единицей скелетной мышцы является
мышечное волокно – многоядерная клетка, образовавшаяся из мезодермы слиянием нескольких сотен, а иногда и тысяч клеток. Длина мышечного волокна достигает 100–1000 мкм, толщина 20–80 мкм Покрыто мышечное волокно двухслойной мембраной – сарколеммой. Непосредственно к цитоплазме прилежит плазмалемма – обычная плазматическая мембрана толщиной 7,5 нм. Через некоторое пространство, заполненное соединительнотканными волокнами, располагается базальная мембрана (5,5 нм), которая является общей для пучка из 20–40 мышечных волокон.
Основное внутриклеточное пространство мышечного волокна заполнено миофибриллами диаметром 1–2 мкм. Их около 2000 и они тянутся вдоль всего мышечного волокна и сформированы в саркомеры.
Саркомер скелетных мышц. Особенностью скелетной, как и всех поперечно-полосатых мышц, является поперечная исчерченность внутреннего содержимого цитоплазмы характерными структурами саркомерами (Рис. ).
149

Рис. 54. Саркомер скелетных мышц.
Длина саркомера (от Z-пластины до Z-пластины) в скелетных мышцах составляет 2,2 мкм. В этих структурах располагаются основные сократительные белки актин и миозин. Строгая упорядоченность их в саркомерах приводит с чередованию оптически более плотных и менее плотных структур.
К Z-пластинам саркомера симметрично по обе стороны прикрепляются нити актина. Между ними в оптически менее плотной (изотропной) зоне I-дисков расположены нити миозина. Посредине каждого I-диска имеется М-полоса – особая мембрана, на которой фиксируются нити миозина. Частично нити актина и миозина перекрываются, образуя оптически более плотную (анизотропную) зону или А-диск. Светлую часть А-диска Н-полосу, содержащую только нити актина, посередине пересекает Z-пластина.
Триада скелетных мышц представляет собой совокупность структур, обеспечивающих запуск сокращения в ответ на раздражение сарколеммы. Она образована тремя структурами (см. рис):
1.Т-системой – впячивания плазматической мембраны внутрь мышечного волокна диаметром около 0,03 мкм.
2.Концевыми цистернами саркоплазматического ретикулума (СПР).
3.Продольными каналами СПР.
Обычно триада располагается вблизи Z-пластин саркомера.
Структура и функция сократительных белков
Основную сократительную функцию во всех видах мышц осуществляют тонкие и толстые нити-миофиламенты (миофибриллы) актин и миозин.
Вспомогательную – регуляторную осуществляют тропомиозин (TrM, ММ:68 кD) и комплекс тропонина (Tr, ММ:70 кD), который состоит из субединиц (Рис. 55)
150