

Класс мхи
.docЖизненный цикл высшего растения состоит из двух фаз (или поколений) - бесполой, или спорофазы (спорофита), и половой, или гаметофазы (гаметофита). Началом развития полового поколения является спора, которая, как нам уже известно, образуется внутри спорангии. В спорангии может образовываться самое различное число спор. У равноспоровых высших растений, у которых споры еще не дифференцированы на микроспоры и мегаспоры, в каждом спорангии обычно образуется не менее 32 спор и лишь очень редко только 8 спор.
У разноспоровых высших растений число микроспор в каждом микроспорангии также обычно не бывает меньше 32, но в мегаспорангии, как правило, образуется только одна мегаспора.
Развитие спор внутри спорангии, особенно развитие мегаспор, происходит довольно сложным путем. Внутри спорангии закладывается первичная спорообразующая ткань. В результате нескольких митотических делений всех спорогенные клетки или только часть их дают начало материнским клеткам спор, или спороцитам. Из каждого спороцита в результате мейоза может образовываться 4 споры. Число хромосом в спорах по сравнению со спороцитом в двое меньше, что объясняется редукцией числа хромосом, происходящей в процессе мейоза следовательно, именно здесь происходит смена ядерных фаз, переход спорофазы в гамето фазу.
Микроспоры и мегаспоры окружены оболочкой (спородермой). Спородерма выполняет разнообразные функции, связанные с жизнедеятельностью споры. На ранних стадиях развития оболочка обеспечивает избирательное поступление веществ из полости спорангия (или гнезда пыльника) в развивающуюся спору. Покидая материнское растение, споры некоторое время существуют самостоятельно. В этих условиях на их оболочку, не прикрытую другими клетками, ложится полная ответственность за сохранение содержимого от высыхания, чрезмерного увлажнения или механических повреждений. Кроме этого, являясь наружным скелетом, спородерма сохраняет определенную форму.
Большое количество жизненно важных функций, ложащихся на оболочку всего лишь одной клетки, вызвало необходимость постоянного совершенствовать строение спородермы. В результате выработалась чрезвычайно прочная, устойчивая к высоким температурам, химическим реагентам и воздействию микроорганизмов, совершенная по конструкции, форме и скульптуре оболочка. Особенностью наружных слоев спородермы является также их поистине удивительная сохранность в ископаемом состоянии.
Существенным признаком в строении спородермы являются также ее форма, определяющаяся многими факторами: соединением спор в тетраде, расположением апертур, свойствами каллозы и т. д. Для бессеменных растений наиболее характерны споры, имеющие тетраэдрическую (плауны, многие папоротники, мхи) или бобовидную, переходящую в эллипсовидную (многие папоротники) форму. Хвощи и большинство моховидных имеют шарообразные споры. Голосеменные унаследовали эллипсовидную и шарообразную формы, в некоторых линиях эволюции трансформировав ее путем образования воздушных мешков. Спородерма современных растений составлена из нескольких хорошо различимых слоев, имеющих различное происхождение и строение. Самый наружный слой - перина - характерен для многих спор мхов, папоротников, хвощей. В виде фрагментов образуется у некоторых голосеменных и, как исключение, представляет самый верхний слой спородермы некоторых очень примитивных цветковых растений. Он формируется из структур, образующихся в клетках стенках спорангия или гнезда пыльника. Следующий слои спородерма - экзина и интина, образуются протопластом самой споры.
Морфологические признаки слоев спородермы формируются постепенно в процессе развития спор. Сразу же после мейоза вокруг каждой споры образуются временная оболочка – каллоза.
Содержимое споры представляет собой первую клетку гаметофазы, и из нее в результате ряда митотических делений развивается гаметофит. Каждый изоспора дает начало обоеполому многоклеточному гаметофиту, на котором развиваются многоклеточные половые органы - антеридии и архегонии. Из гетероспор также развиваются гаметофиты, но содержимое микроспор дает начало только мужским гаметофитам, а содержимое мегаспор - женским гаметофитам.
В процессе исторического развития от псилофитов произошли более сложные растения - моховидные. Отдел моховидных включает в себя разнообразные виды мхов, которых насчитывается около 25 тыс. Эти растения широко распространены по земному шару. В умеренной и холодной зонах северного полушария растут зеленые мхи, среди которых широко распространен кукушкин лен. Он образует рыхлые дернины на кочках в сырых хвойных лесах. На болотах, заболоченных лугах, во влажных лесах растут сфагновые мхи. Мхи, как правило, многолетние и в основном низкорослые растения, "лилипуты" растительного мира. Высота их чаще всего не превышает 30 см.
Среди ныне живущих высших растений мхи имеют наиболее примитивное строение. Так, мох маршанция не имеет четко выраженных вегетативных органов, его тело не расчленено на стебель и листья и имеет вид зеленой пластинки. Большинство же мхов листостебельные растения (правда, так называть их можно лишь условно, так как листовидные образования мхов не гомологичны настоящим листьям, но аналогичны по функциям). У некоторых мхов, например у зеленых, имеются ризоиды - тонкие нити, состоящие из одного ряда клеток. Другие мхи (например, сфагнум) не имеют ризоидов. Ризоидами мхи укрепляются в почве и поглощают из нее воду и минеральные вещества.
Внутреннее строение мхов сравнительно простое. У них слабо развита проводящая система, а передвижение воды, минеральных и органических веществ происходит по особым вытянутым в длину клеткам, расположенным друг над другом. По мнению ряда ученых, недоразвитость проводящей системы ограничивает рост мха в высоту. В то же время на поперечном срезе под микроскопом можно увидеть разнообразные ткани стебля: кору, основную ткань, центральный пучок.
Листья у мха простые, расположены на стебле спирально. У кукушкина льна листья зеленые, а у сфагнума светло-зеленые. Рассматривая под микроскопом строение листа сфагнума, можно различить два типа клеток: одни узкие, живые, заполненные хлоропластами, а другие - крупные, бесцветные, лишенные хлоропластов мертвые клетки, их называют водоносными, они окружают клетки с хлоропластами. В клетках с хлоропластами происходит фотосинтез. Благодаря водоносным клеткам листья сфагнума обладают гигроскопическим свойством, т. е. способны удерживать воду.
Размножаются мхи как половым, так и бесполым способом. При этом происходит чередование поколений: бесполого (спорофит) и полового (гаметофит). Развитие кукушкина льна, например, происходит следующим образом. На верхней части стебля мха в коробочках развиваются споры. Попадая на почву, спора прорастает тонкой зеленой нитью (протонема), которая сходна по строению с нитчатыми водорослями. (Это сходство свидетельствует о родстве мхов с водорослями и происхождении первых наземных растений от древних водорослей.) Нить ветвится, на ней появляются почки, из которых затем вырастают стебли с листьями - взрослые растения мха. Кукушкин лен - двудомное растение (многие мхи однодомные или многодомные). На мужских растениях развиваются мужские половые органы антеридии, в которых образуются сперматозоиды, а на женских растениях - женские половые органы архегонии, в которых созревают яйцеклетки. Во время дождя, обильной росы с каплями воды сперматозоиды попадают в архегонии и сливаются с яйцеклетками. Из оплодотворенной яйцеклетки - зиготы - развиваются коробочка со спорами. Таким образом, взрослое растение мха - половое поколение (гаметофит), а коробочка со спорами - бесполое поколение (спорофит).
Как и другие растения, мхи участвуют в круговороте веществ в природе: синтезируют органические вещества и выделяют кислород в атмосферу. Мхи устойчивы к резким колебаниям температуры, к избыточному увлажнению и засухе, живут на бедных почвах, скалах, камнях, образуют сообщества в таких местах, где другие высшие растения жить не могут. Постепенно отмирая, мхи обогащают почву органическими веществами, подготавливают ее для заселения другими растениями.
Велика роль мхов в тундре и лесотундре, где они защищают вечную мерзлоту от нагревания солнечными лучами. Нарушение целостности мохового покрова (при пожарах, строительстве, проезде транспорта и др.) ведет к увеличению глубины протаивания и к образованию оврагов, котловин, оползней и др.
Большую роль играют сфагновые мхи в формировании и жизни болот. Из отмерших остатков сфагнума образуются залежи торфа. Сфагнум растет верхушкой стебля, а его нижняя часть отмирает ("отторфовывается"). Так в течение многих лет образуются огромные залежи торфа. Процесс торфообразования происходит благодаря застойному переувлажнению, отсутствию кислорода и созданию мхами кислой среды. Эти условия оказываются неблагоприятными для процессов гниения, что препятствует разложению сфагнума. Торф - ценное сырье для получения воска, парафина, аммиака, спирта и т.д. Он используется в медицине, в строительной промышленности, в качестве топлива, удобрений. Значение болот однако не сводится только к образованию в них торфа. На болотах формируется своеобразное сообщество, в котором значительное место занимают клюква, лекарственные растения, обитают животные, в том числе и промысловые. Но главное, болото - это резервуар пресной воды, он, как губка, впитывает в себя все осадки, которые затем вытекают из него в виде ручейков и впадают в реки. Поэтому не всегда можно осушать небольшие болота, так как это может вызвать обмеление и заиление рек, наступление засухи.
Многие мхи угнетают рост и жизнедеятельность таких микроорганизмов, как кишечная палочка, холерный вибрион, золотистый стафилококк, сальмонелла. Эта способность мхов угнетать, подавлять жизнедеятельность других организмов является средством их самозащиты, приспособлением к выживанию в условиях острой борьбы за существование.
КЛАСС МХИ, или ЛИСТОСТЕБЕЛЬНЫЕ МХИ (MUSCI), или БРИОПСИДЫ (BRYOPSIDA)
Это наиболее примитивные из групп наземных растений. Они мало приспособлены к жизни на суше, поэтому привязаны к влажным, затененным местам. У мхов нет настоящей сосудистой ткани (ксилемы и флоэмы), нет настоящих корней (вместо них нитчатые выросты стебля - ризоиды). Вода и минеральные соли поглощаются всей поверхностью тела, в том числе и ризоидами.
Жизненный цикл мхов включает гаплоидный гаметофит и диплоидный спорофит. Доминирующим поколением является гаплоидный гаметофит, который принимает на себя функции фотосинтеза, водоснабжения и минерального питания. Для обеспечения полового процесса необходима капельножидкая среда. Половое и бесполое поколение мхов не разделены, а представляет одно растение. Гаметофит развивается из гаплоидной споры. У разных видов мхов гаметофит может быть однополым (двудомным) и разнополым (однодомным). Органы полового размножения (гаметангии) образуют подвижные сперматозоиды и неподвижные яйцеклетки. Оплодотворение яйцеклетки происходит внутри женского полового органа. Из зиготы медленно развивается диплоидный спорофит, который представляет собой коробочку (спорангий), находящуюся на гаметофите и получающую от него питание. В коробочке путем мейоза образуются гаплоидные споры.
Наиболее известными представителями мхов являются кукушкцнлен и сфагнум. Стебли сфагнума имеют,светло-зеленый цвет и несут на себе мутовки ветвей, листья которых не имеют жилкования. У взрослых растений сфагнума нет ризоидов, и на болотах они образуют плотные подушки из прямостоячих ветвей. Листья (филлоиды) наряду с хлорофиллосодержащими клетками имеют мертвые клетки с утолщенными стенками, впитывающие воду. Сфагновые мхи - торфообразователи. Торф образуется в результате накопления и уплотнения отмерших нижних частей гаметофита. Их разложение не происходит из-за низкой кислотности и недостатка кислорода. Торф широко используется как топливо, удобрение и сырье в промышленности. Сфагнум обладает бактерицидными свойствами, и его используют в медицине.
Самый крупный класс мохообразных, насчитывающий около 800 родов и до 15 тысяч видов, относимых примерно к 80 семействам.
Все мхи — листостебельные растения, но в отличие от листостебельных юнгерманниевых мхи по большей части обладают хорошо выраженной нитчатой протонемой, все листья у мхов нормально развиты (нет амфигастриев) и всегда цельные, а коробочка спорогония обычно имеет колонку и никогда не содержит элатер.
Класс мхов разделяют на три подкласса, хорошо различающихся между собой по строению спорогония и некоторым особенностям гаметофита: Сфагновые (Sphagnidae), Андреэевые (Andreaeidae) и Бриевые (Bryidae); первые два класса невелики по объему и содержат соответственно около 300 и около 100 видов.
Подкласс Сфагновые (Sphagnidae).
Все представители подкласса сфагновых мхов — влаголюбивые растения, и побег сфагнов обладает многочисленными особенностями, позволяющими растению удерживать и сохранять воду.
Стебель сфагнов прямостоячий, редкооблиственный, с многочисленными густооблиственными боковыми ветвями (см. например, изображения сфагна дубравного или сфагна Вульфа).
Ветви на верхушке стебля скучены в плотную головку, ниже — собраны в пучки; при этом часть ветвей каждого пучка оттопырена в сторону, благодаря чему побеги соединяются в более или менее плотные дерновинки или подушечки. Другая часть ветвей каждого пучка свисает вдоль стебля, образуя своеобразный фитиль, по которому передвигается вода. Ризоидов у взрослого растения нет. Побег нарастает в верхней части и постепенно отмирает снизу. Некоторые ветви на верхушке побега растут быстрее остальных и при отмирании стебля снизу дают начало новому побегу, вследствие чего подушечки сфагна постепенно увеличиваются в объеме.
Снаружи стебель главного побега и ветвей (рис. 8) покрыт бесцветной кожицей — гиалодермисом, состоящим из крупных мертвых клеток, в стенках которых у многих видов имеются отверстия — поры. В гиалодермисе ветвей часто встречаются водосборные ретортовидные клетки с отверстиями на концах.
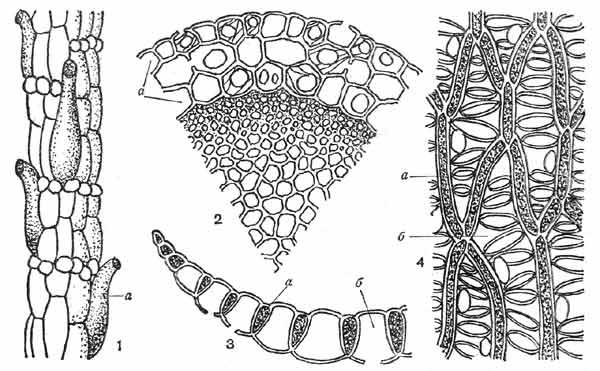
Рис. 8. Строение стебля и листа сфагна:
1 — поверхность стебля с ретортовидными клетками (а), 2 — поперечный срез стебля (а — гиалодермис), 3 — поперечный срез листа, 4 — лист с поверхности (а — фотосинтезирующие клетки, б — гиалиновые клетки)
Листья сфагнов однослойные, без жилки, разнообразной формы; листья главного побега обычно отличаются по форме, микроструктуре и размерам от веточных листьев того же вида (см. иллюстрации листьев мхов рода Сфагнум — Sphagnum).
Пластинка листа (рис. 8) состоит из клеток двух типов — узких живых фотосинтезирующих клеток, оболочка которых нередко пропитана красящими веществами, и широких мертвых водоносных клеток. Прозрачные, или гиалиновые, мертвые клетки, как и клетки гиалодермиса стебля, имеют отверстия — поры. Микроскопические особенности строения листьев играют важную роль при определении видов сфагновых мхов. Через поры гиалиновых клеток листьев и стебля, через отверстия ретортовидных клеток, а также по капиллярным промежуткам между свисающими ветвями и стеблем вода легко передается от одной части растения к другой, и это объясняет замечательную способность сфагнов быстро и в большом количестве впитывать воду. При испарении воды гиалиновые клетки заполняются воздухом, поэтому сухие растения сфагна приобретают беловатый оттенок.
Антеридии и архегонии у сфагнов располагаются на особых ветвях (рис. 9, 4) одного и того же растения или на разных растениях. Антеридии сидят в пазухах покровных листьев. Ветви с антеридиями обычно булавовидно вздуты и ярко окрашены в различные цвета, но после созревания антеридиев ветви удлиняются и не отличаются по окраске от остальных отстоящих ветвей растения. Архегонии располагаются на концах укороченных почковидных женских веточек также под защитой покровных листьев.

Рис. 9. Строение сфагновых мхов:
1 — спора, 2 — молодая протонема, 3 — пластинчатая зрелая протонема с молодым листостебельным побегом (гаметофором), 4 — часть стебля с антеридиальными (а) и с архегониальными (б) ветвями, 5 — антеридиальная ветвь, частично лишенная листьев (а — антеридий), 6 — вскрывающийся антеридий, 7 — сперматозоид, 8 — верхушка архегониальной ветви на продольном срезе, 9 — продольный срез спорогония (а — ложноножка, б — стопа, в — колонка, г — спорангий, 9 — колпачок), 10 — зрелый спорогоний на ложноножке (а — ложноножка, б — остатки колпачка, в — коробочка, г — крышечка).
Молодой спорогоний (рис. 9, 4), находящийся внутри колпачка, прикрыт снаружи покровными листьями и состоит из стопы и коробочки; ножка у спорогония сфагновых мхов недоразвита. Со дна коробочки поднимается куполообразная колонка, прикрытая сводообразным спорангием. При созревании коробочка прорывает колпачок, который остается в виде оборки при ее основании, и в это же время часть стебля, расположенная между стопой и покровными листьями, сильно удлиняется, образуя ложноножку, которая выносит шаровидную коробочку далеко за пределы покровных листьев (рис. 9, 10). Коробочка обычно вскрывается путем резкого отбрасывания крышечки, при этом споры выбрасываются на расстояние до 10 см. Процесс вскрывания коробочки и выбрасывания спор легко проследить, подсушивая выбранные из влажной дернинки растения со зрелыми спорогониями.
Протонема имеет сначала вид короткой нити, а затем лопастной пластинки (рис. 9, 2-3), на которой закладывается почка, дающая впоследствии листостебельный побег, или гаметофор.
Подкласс Андреэевые (Andreaeidae).
Представители подкласса андреэевых мхов отличаются от остальных мхов целым рядом признаков. Спора андреэевых, прорастая, дает начало стелющейся по поверхности субстрата (обычно камней и скал) разветвленной пластинчатой протонеме с листообразными придатками и древовидными выростами (рис. 10). На протонеме появляются почки, из которых вырастает гаметофор, прикрепляющийся к субстрату толстыми цилиндрическими или пластинчатыми ризоидами. Спорогоний андреэевых сидит на верхушке стебля; он обладает колонкой, не доходящей до верхушки коробочки, и не имеет развитой ножки. Созревшую коробочку за пределы покровных листьев выносит удлиняющаяся ложноножка. Коробочка вскрывается 4-8 продольными щелями (рис. 15, 1).
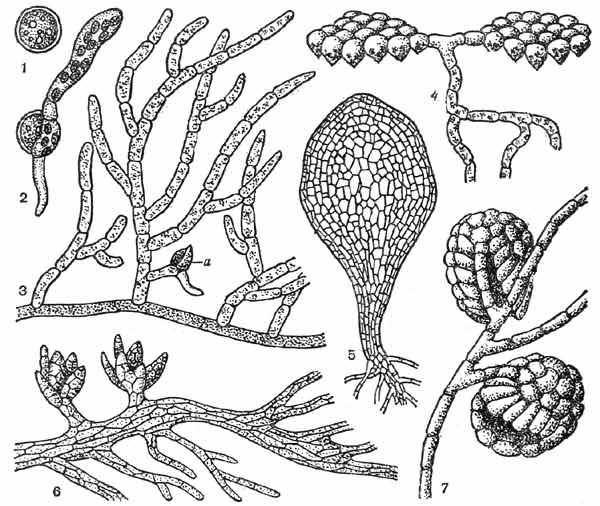
Рис. 10. Спора и протонема аидреэевых и бриевых мхов:
1 — спора у фунарии, 2 — прорастающая спора у фунарии, 3 — нитчатая протонема с почечкой (а) у фунарии, 4 — протонема схистостеги, 5 — пластинчатая протонема тетрафиса, 6 — протонема андреэи, 7 — протонема и мужской гаметофор у буксбаумии
Подкласс Бриевые, или Зеленые Мхи (Bryidae).
Подкласс бриевых, или зеленых, мхов — наиболее многочисленная группа мохообразных. Протонема этих мхов обычно имеет вид сильноразветвленной, стелющейся по поверхности субстрата нити, от которой в почву отходят незеленые ризоиды (рис. 10, 2-3).
У некоторых бриевых протонема листовидная, например у тетрафиса (Tetraphis)(рис. 10, 5). У схистостеги (Schistostega) — мха, растущего в расщелинах скал, в пещерах, дуплах деревьев, одни ветви протонемы, прикрепляющие ее к субстрату, нитчатые, другие, располагающиеся над субстратом, имеют вид однослойных пластинок, направленных перпендикулярно к лучам света (рис. 10, 4). Пластинки эти состоят из линзовидно-воронковидных клеток, в воронковидной части которых находятся зеленые пластиды (хлоропласты), а над хлоропластами, подобно хрусталику в глазном яблоке, располагается бесцветная масса цитоплазмы. Рассеянные лучи света, падающие на поверхность пластинки, преломляются в линзовидной части клетки и фокусируются на хлоропластах; при этом часть лучей отражается, что вызывает свечение пластинки зеленым светом (отсюда народное название схистостеги — «светящийся мох»).
Протонема некоторых бриевых мхов (например, фаска — Phascum, поттии — Pottia) в противоположность листостебельному побегу этих растений многолетняя, зимующая. У буксбаумии (Buxbaumia) протонема также многолетняя, а мужской гаметофор у буксбаумии сведен до одного листа, под защитой которого находится антеридий (рис. 10, 7).
Из почек, образующихся на протонеме, вырастают листосте-бельные побеги, длина которых варьирует от долей миллиметра до полуметра. Побеги бывают простыми или разветвленными, прямостоячими, приподнимающимися или восходящими, висячими, стелющимися или лежачими (ползучими).
Способ ветвления мхов часто связан с расположением половых органов, а точнее, спорогониев. У мхов, у которых спорогоний располагается на верхушке главного побега или столь же мощных ветвей (верхоплодные мхи), побеги простые либо вильчато или мутовчато ветвящиеся. У мхов, у которых спорогоний располагается на верхушке сильно укороченных почковидных боковых ветвей (бокоплодные мхи), побеги ветвятся обычно по-иному (главным образом перисто) и обладают хорошо выраженным главным стеблем. У некоторых бриевых мхов (например, климация) имеются мощные подземные корневищеподобные побеги (см. например, изображения климация древовидного или леукодона беличьего), которые служат для вегетативного размножения и, выйдя на поверхность, превращаются в обычные побеги.
Стебель у бриевых мхов тонкий, 0,1-1 мм в диаметре, почти бесцветный либо окрашенный — зеленоватый, красноватый, бурый или черноватый, в нижней части или почти на всем своем протяжении покрытый ризоидами — однорядными многоклеточными, обычно бурыми или красноватыми, простыми или разветвленными волосками (рис. 11, 1). Ризоиды, соприкасающиеся с субстратом, внедряются в него, прикрепляют к субстрату растение и снабжают его водой с растворенными в ней минеральными солями; на остальной части стебля ризоиды нередко образуют густой войлок, который может объединять отдельные побеги в более или менее плотную дерновинку.

Рис. 11. Строение бриевых мхов:
1 — стебель на поперечном срезе (на поверхности стебля видны ризоиды, в центре — проводящий пучок), 2 — парафилл, 3 — корзиночка с выводковыми телами у тетрафиса, 4 — ложноножка с выводковыми телами у аулакомний, 5 — чашевидная (а) и проросшая (б) мужская почка погоната, 6 — обоеполая почка брия на продольном разрезе (а — антеридий, б — архегоний, в — парафиза), 7 —мужские растения сплахна, 8 — спорогоний сплахна с зонтиковидной апофизой
Листья бриевых мхов не имеют черешка и прикрепляются к стеблю более или менее широким основанием всегда цельной пластинки поперек, наискось или почти вдоль стебля, располагаясь в два или несколько вертикальных рядов. Побеги могут быть густо- и рыхлооблиственными. Иногда у верхушки стебля листья более тесно скучены, чем на остальном его протяжении, и образуют розетку или хохолок. Во влажном состоянии листья плотно прижаты к стеблю, черепитчато налегая один на другой, или отстоят от стебля под тем или иным углом.
В зависимости от величины угла различают листья прямостоячие (угол до 15°), прямоотстоящие (угол до 30°), отклоненные (угол около 45°), оттопыренные (угол 45-90°) и отогнутые (угол более 90°); иногда основание листа может быть прямоотстоящим, а верхушка — отогнутой. Обычно листья равномерно обращены во все стороны, реже — одностороннеобращенные. У сухих растений по сравнению с влажными характер отхождения листьев от стебля может меняться, а пластинка листа иногда завивается вокруг стебля либо изгибается или закручивается.
Пластинка обычного фотосинтезирующего листа бывает плоской, ложковидно- или чашевидно-вогнутой, килеватой, желобчатой, трубчатой, продольно-складчатой (далее в тексте просто складчатой) или поперечно-волнистой (далее в тексте просто волнистой).
По очертаниям различают листья треугольные, сердцевидные, округлые, яйцевидные, эллиптические, продолговатые, языковидные, лопатчатые, ланцетные, линейные, серповидные, крючковидные, треугольно-сердцевидные, продолговато-ланцетные и т. д. Пластинка листа (за исключением жилки, а иногда и каймы) всегда однослойная и состоит из клеток, которые могут различаться между собой по форме, величине и строению. Основание листа бывает суженным или расширенным, закругленным или выемчатым; иногда основание низбегает по стеблю или, разрастаясь, образует более или менее крупное, охватывающее стебель влагалище.
Клетки, расположенные в углах основания пластинки, могут отличаться по форме, величине и цвету от остальных клеток пластинки, иногда образуя служащие для сохранения воды участки — листовые крылья; если крылья сильно вздуты, они называются листовыми ушками (в описаниях те и другие обозначены как ушки).
Края листовой пластинки бывают цельными или зубчатыми; они также могут быть плоскими, либо загнутыми (завороченными) на верхнюю сторону, либо отогнутыми (отвороченными) на нижнюю сторону. Клетки, расположенные по краю и отличающиеся от остальных клеток пластинки формой, величиной и окраской, образуют плоскую или вздутую кайму. Верхушка листовой пластинки может быть выемчатой, закругленной, тупой, длинно-или короткозаостренной, шиловидной, прямой или крючковато-изогнутой, колпачковидной, с остроконечием.
У большинства видов бриевых мхов листья обладают более или менее мощной жилкой, которая представляет собой тяж, сложенный несколькими слоями клеток. Жилка бывает узкой или широкой, на верхней стороне жилки иногда располагаются продольные зеленые пластинчатые выросты — ассимиляционные пластиночки, или ассимиляторы. Жилка чаще простая (но может также быть вильчатой или двойной), короткая либо длинная, заканчивающаяся в нижней или верхней половине листа. Она может выступать из верхушки листа в виде острия либо волоска.
Микроскопические особенности строения пластинки обычно играют решающую роль в определении видов бриевых мхов.
Фотосинтезирующие листья на протяжении одного и того же стебля или листья главного побега и боковых ветвей, а также спинные, боковые и брюшные листья у плоскооблиственных побегов могут различаться между собой по форме и величине. Листья на подземных или стелющихся по поверхности субстрата корневище-подобных частях побега по форме, величине и строению обычно отличаются от нормальных фотосинтезирующих листьев и часто имеют вид чешуи. Быстрорастущие побеги с расставленными чешуевидными листьями называются столоновидными побегами. Нередко от обычных вегетативных листьев отличаются по форме и величине, а иногда и по окраске листья, окружающие и защищающие половые органы, расположенные на верхушках побегов и ветвей (рис. 11, 5). Листья, окружающие антеридии, называются перигониальными, а окружающие архегонии — перихециальными.