

Глава 4. Основные характеристики мышечной деятельности
4.1. Скелетные мышцы.
Скелетные (поперечнополосатые) мышцы — это "машины", преобразующие химическую энергию непосредственно в механическую и тепловую. Сокращение мышц возникает в ответ на электрические импульсы, приходящие к ним от а- мотонейронов — нервных клеток, лежащих в передних рогах спинного мозга. Мышцы и иннер-вирующие их мотонейроны составляют нервно-мышечный аппарат человека. В результате сократительной деятельности скелетных мышц осуществляется поддержание позы человека, перемещение частей тела относительно друг друга, передвижение человека в пространстве.
Основным морфо-функциональным элементом нервно-мышечного аппарата является двигательная единица (ДЕ). ДЕ — это мотонейрон с иннервируемыми им мышечными волокнами. Аксон мотонейрона из спинного мозга проходит в составе периферических нервов до мышцы, внутри которой разветвляется на множество концевых веточек. Каждая концевая веточка заканчивается на одном мышечном волокне, образуя нервно-мышечный синапс. Импульсы, идущие по аксону мотонейрона, активируют все иннервируемые им мышечные волокна. Поэтому ДЕ функционирует как единое морфофунк-циональное образование.
Скелетная мышца состоит из пучков вытянутых в длину клеток — мышечных волокон, обладающих тремя свойствами: возбудимостью, проводимостью и сократимостью (см.главу I). Отличительной чертой мышечных клеток от клеток, не обладающих свойством сократимости, является наличие саркоплазматического ретикулума. Он представляет собой замкнутую систему внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу. В мембране саркоплазматического ретикулума находятся две транспортные системы, обеспечивающие освобождение от ретикулума ионов кальция при возбуждении и их возврат из миоплазмы обратно в ретикулум при расслаблении мышцы. В механизме освобождения ионов кальция из ретикулума при возбуждении мышечной клетки важную роль играет система поперечных трубочек (Т-система), представляющих собой впячивания поверхностной мембраны мышечного волокна. К противоположным сторонам поперечной трубочки примыкают боковые цистерны ретикулума. Две терминальные цистерны ретикулума вместе с трубочкой образуют так называемую триаду — анатомическую
146
структуру, в зоне которой нервные импульсы, распространяющиеся по поперечным трубочкам вглубь мышечного волокна, запускают процесс выхода ионов кальция из саркоплазматического ретикулума и, следовательно, всю последующую цепочку изменений, приводящую, в конечном итоге, к развитию сокращения мышцы.
Мышечные волокна имеют диаметр от 10 до 100 мкм и длину от 5 до 400 мм (в зависимости от длины мышцы). В каждом мышечном волокне содержится до 1000 и более сократительных элементов ми-офибрилл, толщиной 1-3 мкм. Каждая миофибрилла состоит из множества параллельно лежащих толстых и тонких нитей — миофиламен-тов. Толстые нити состоят из молекул белка миозина, а тонкие — из белка актина. Миозиновые нити имеют отходящие от них биполярно поперечные выступы около 20 нм, с головками, состоящими примерно из 150 молекул миозина. Во время сокращения каждая головка миозина, или поперечный мостик, может связывать миозиновую нить с соседней актиновой. Кроме того, в состав тонких нитей входят еще два белка — тропонин и тропомиозин, необходимые для развития процессов сокращения и расслабления мышцы.
Расположение миозиновых и тонких актиновых белковых нитей строго упорядочено (рис.4.1.). Пучок лежаших в середине саркомера нитей миозина выглядит в световом микроскопе как темная полоска. Благодаря свойству двойного лучепреломления в поляризованном свете (то есть анизотропии) она называется А-диском. По обе стороны от А-диска находятся участки, которые содержат только тонкие нити актина и поэтому выглядят светлыми. Эти изотропные J-диски тянутся до Z-пластин. Благодаря такому периодическому чередованию светлых и темных полос миофибриллы скелетной мышцы
Р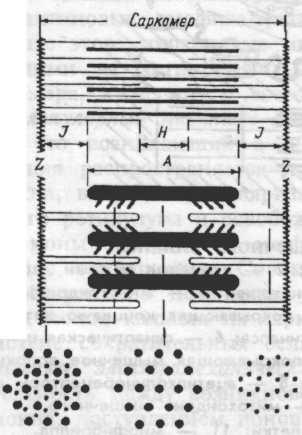 ис.4.1.
Схема саркомера мышечного волокна
и взаимного расположения
толстых миозиновых
и тонких актиновых миофиламентов.
ис.4.1.
Схема саркомера мышечного волокна
и взаимного расположения
толстых миозиновых
и тонких актиновых миофиламентов.
Поперечный срез миофибриллы дает
представление о гексагональном
распределении актиновых и миозиновых
нитей.
Z — линии, разделяющие два соседних
саркомера;
J — изотропный диск;
А — анизотропный диск;
Н — участок с уменьшенной
анизотропностью.

 выглядят
исчерченными (поперечно-полосатыми).
Если
мышца расслаблена,
то в средней части А-диска различается
менее плотная Н-
зона, состоящая только из толстых
миофиламентов. Н- зона не просматривается
во время сокращения мышцы. По середине
J-дис-ка
проходит темная полоска — это Z
линия. Участок миофибриллы между
двумя Z
линиями называется саркомером.
выглядят
исчерченными (поперечно-полосатыми).
Если
мышца расслаблена,
то в средней части А-диска различается
менее плотная Н-
зона, состоящая только из толстых
миофиламентов. Н- зона не просматривается
во время сокращения мышцы. По середине
J-дис-ка
проходит темная полоска — это Z
линия. Участок миофибриллы между
двумя Z
линиями называется саркомером.
Передача возбуждения в нервно-мышечном синапсе. Структура нервно- мышечного синапса представлена на рисунке 4.2. В процессе передачи возбуждения с нерва на мышечные волокна выделяют три последовательных процесса: 1. электрический, включащий достижение нервным импульсом концевой веточки аксона, деполяризацию и повышение проницаемости ее мембраны, выделение аце-тилхолина (АХ) в синаптическую щель; 2. химический, основу которого составляет диффузия медиатора АХ к постсинаптической мембране и образование на ней его комплекса с холинорецептором; 3. электрический, включащий увеличение ионной проницаемости постсинаптической мембраны, возникновение локального электри-
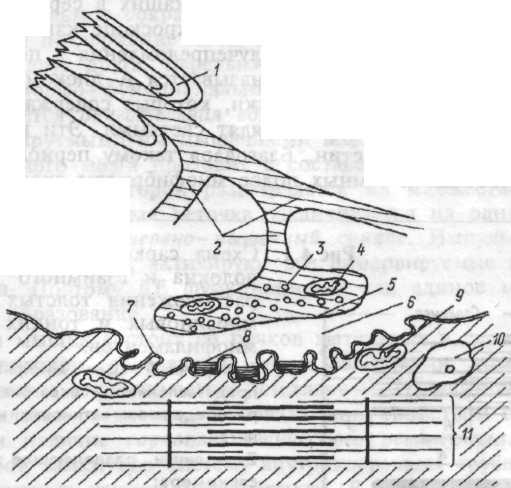
Рис.4.2. Схема элементов нервно- мышечного синапса.
1 — миэлиновая оболочка аксона; 2 — концевые веточки аксона; 3 — пузырьки, содержащие ацетилхолин; 4 — митохондрия; 5 — пресинаптическая мембрана, покрывающая концевую веточку аксона в зоне нервно-мышечного синапса; 6 — синаптическая щель; 7 —постсинаптическая мембрана, покрывающая мышечное волокно в зоне нервно-мышечного синапса; 8 — ацетилхолинорецепторы на постсинаптической мембране; 9 — митохондрия мышечного волокна; 10 — ядро мышечной клетки; 11 — миофибрилла.
148
 ческого
потенциала (потенциала
концевой пластинки; ПКП),
развитие
потенциала действия мышечного
волокна.
ческого
потенциала (потенциала
концевой пластинки; ПКП),
развитие
потенциала действия мышечного
волокна.
Запасов АХ в нервном окончании достаточно для проведения лишь примерно 10 000 импульсов. При длительной же импульсации мотонейрона, несмотря на постоянный синтез АХ (см. главу 3), его содержание в концевых веточках может постепенно уменьшаться. В результате этого возможны нарушения передачи возбуждения в нервно-мышечных синапсах — пресинаптический нервно-мышечный блок.
Временно возникающий на постсинаптической мембране комплекс "АХ- рецептор" после прохождения каждого импульса разрушается ферментом ацетил-холинэстеразой. Однако при длительной высокочастотной импульсации мотонейрона (например при длительной и напряженной мышечной работе) АХ не успевает разрушаться и накапливается в синаптической щели. Способность постсинаптической мембраны к генерации ПКП при этом снижается и развивается частичный или полный постсинаптический нервно- мышечный блок, приводящий либо к частичному, либо даже полному прекращению развития потенциалов действия на мембране мышечного волокна.
Механизмы сокращения мышечного волокна. В покоящихся мышечных волокнах при отсутствии импульсации мотонейрона поперечные миозиновые мостики не прикреплены к актиновым ми-офиламентам. Тропомиозин расположен таким образом, что блокирует участки актина, способные взаимодействовать с поперечными мостиками миозина. Тропонин тормозит миозин — АТФ-азную активность и поэтому АТФ не расщепляется. Мышечные волокна находятся в расслабленном состоянии.
При сокращении мышцы длина А-дисков не меняется, J-диски укорачиваются, а Н-зона А-дисков может исчезать (рис. 4.3.). Эти данные явились основой для создания теории, объясняющей сокращение мышцы механизмом скольжения (теорией скольжения) тонких актиновых миофиламентов вдоль толстых миозиновых. В результате этого миозиновые миофиламенты втягиваются между окружающими их актиновыми. Это приводит к укорочению каждого саркомера, а значит, и всего мышечного волокна.
Молекулярный механизм сокращения мышечного волокна состоит в том, что возникающий в области концевой пластинки потенциал действия распространяется по системе поперечных трубочек вглубь волокна, вызывает деполяризацию мембран цистерн саркоплазмати-ческого ретикулума и освобождение из них ионов кальция. Свободные ионы кальция в межфибриллярном пространстве запускают процесс сокращения. Совокупность процессов, обуславливающих распространение потенциала действия вглубь мышечного волокна, выход ионов кальция их саркоплазматического ретикулума, взаимодействие сократительных белков и укорочение мышечного волокна называют "электромеханическим сопряжением". Временная последовательность между возникновением потенциала действия мышечного волокна, поступлением ионов кальция к миофибриллам и развитием сокращения волокна показана на рисунке 4.4.
149




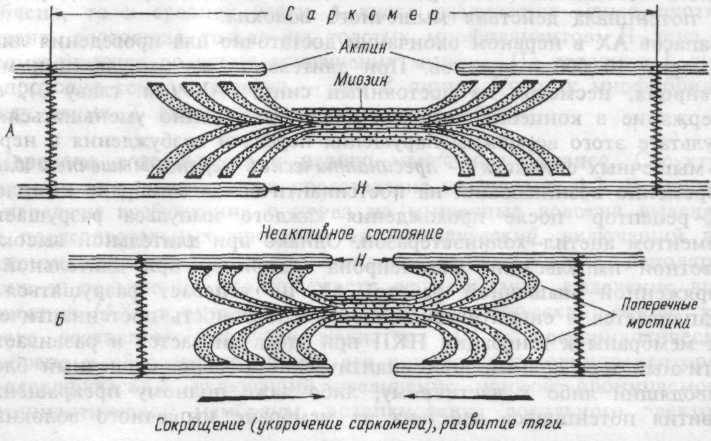
Рис.4.3. Сокращение мышцы.
А. Поперечные мостики между актином и миозином разомкнуты. Мышца находится в расслабленном состоянии.
Б. Замыкание поперечных мостиков между актином и миозином. Совершение головками мостиков гребковых движений по направлению к центру саркомера. Скольжение актиновых нитей вдоль миозиновых, укорочение саркомера, развитие тяги.
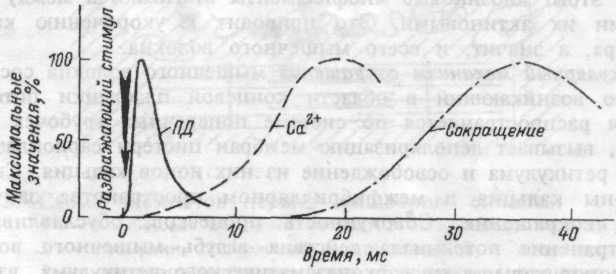
Рис.4.4. Схема временной последовательности развития
потенциала действия (ПД), освобождения ионов кальция (Са2+) и развития изометрического сокращения мышцы.
150
При концентрации ионов Са2+ в межмиофибриллярном пространстве ниже 10" тропомиозин располагается таким образом, что блокирует прикрепление поперечных миозиновых мостиков к нитям актина. Поперечные мостики миозина не взаимодействуют с нитями актина. Продвижение относительно друг друга нитей актина и миозина отсутствует. Поэтому мышечное волокно находится в расслабленном состоянии. При возбуждении волокна Са2+ выходит из цистерн саркоплаз-матического ретикулума и, следовательно, концентрация его вблизи миофибрилл возрастает. Под влиянием активирующих ионов Са2+ молекула тропонина изменяет свою форму таким образом, что выталкивает тропомиозин в желобок между двумя нитями актина, освобождая тем самым участки для прикрепления миозиновых поперечных мостиков к актину. В результате поперечные мостики прикрепляются к актиновым нитям. Поскольку головки миозина совершают "гребко-вые" движения в сторону центра саркомера происходит "втягивание" актиновых миофиламентов в промежутки между толстыми миозиновы-ми нитями и укорочение мышцы.
Источником энергии для сокращения мышечных волокон служит АТФ. С инактивацией тропонина ионами кальция активируются каталитические центры для расщепления АТФ на головках миозина. Фермент миозиновая АТФ-аза гидролизует АТФ, расположенный на головке миозина, что обеспечивает энергией поперечные мостики. Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат используются для последующего ресинтеза АТФ. На миозиновом поперечном мостике образуется новая молекула АТФ. При этом происходит разъединение поперечного мостика с нитью актина. Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока концентрация кальция внутри миофибрилл не снижается до подпороговой величины. Тогда мышечные волокна начинают расслабляться.
При однократном движении поперечных мостиков вдоль актиновых нитей (гребковых движениях) саркомер укорачивается примерно на 1% его длины. Следовательно, для полного изотонического сокращения мышцы необходимо совершить около 50 таких гребковых движений. Только ритмическое прикрепление и отсоединение головок миозина может втянуть нити актина вдоль миозиновых и совершить требуемое укорочение целой мышцы. Напряжение, развиваемое мышечным волокном, зависит от числа одновременно замкнутых поперечных мостиков. Скорость развития напряжения или укорочения волокна определяется частотой замыкания поперечных мостиков, образуемых в единицу времени, то есть скоростью их прикрепления к актиновым миофиламентам. С увеличением скорости укорочения мышцы число одновременно прикрепленных поперечных мостиков в каждый момент времени уменьшается. Этим и можно объяснить уменьшение силы сокращения мышцы с увеличением скорости ее укорочения.
При одиночном сокращении процесс укорочения мышечного волокна заканчивается через 15-50 мс, так как активирующие его ионы кальция возвращаются при помощи кальциевого насоса в
151


 цистерны
саркоплазматического ретикулума.
Происходит расслабление
мышцы.
цистерны
саркоплазматического ретикулума.
Происходит расслабление
мышцы.
Поскольку возврат ионов кальция в цистерны саркоплазматического ретикулума идет против диффузионного градиента, то этот процесс требует затрат энергии. Ее источником служит АТФ. Одна молекула АТФ затрачивается на возврат 2-х ионов кальция из межфибриллярного пространства в цистерны. При снижении содержания ионов кальция до подпорогового уровня (ниже 10 V) молекулы тропонина принимают форму, характерную для состояния покоя. При этом вновь тропомиозин блокирует участки для прикрепления поперечных мостиков к нитям актина. Все это приводит к расслаблению мышцы вплоть до момента прихода очередного потока нервных импульсов, когда описанный выше процесс повторяется. Таким образом, кальций в мышечных волокнах играет роль внутриклеточного посредника, связывающего процессы возбуждения и сокращения.
Режимы и типы мышечных сокращений. Режим сокращений мышечных волокон определяется частотой импульсации мотонейронов. Механический ответ мышечного волокна или отдельной мышцы на однократное их раздражение называется одиночным сокращением. При одиночном сокращении выделяют: 1. фазу развития напряжения или укорочения; 2. фазу расслабления или удлинения (рис.4.5.). Фаза расслабления продолжается примерно в два раза дольше, чем фаза напряжения. Длительность этих фаз зависит от морфофункциональ-ных свойств мышечного волокна: у наиболее быстро сокращающихся волокон глазных мышц фаза напряжения составляет 7-10 мс, а у наиболее медленных волокон камбаловидной мышцы — 50-100 мс.
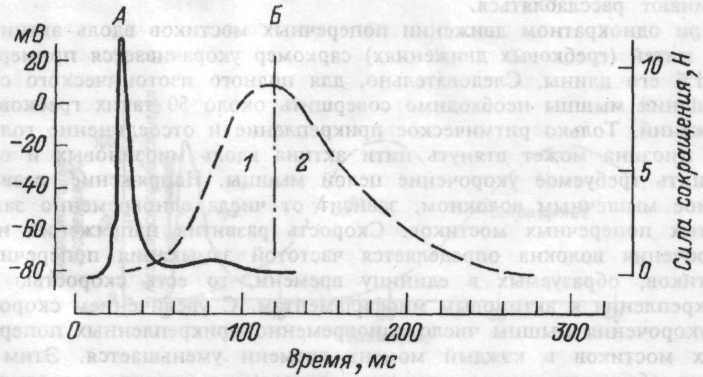
Рис.4.5. Развитие во времени потенциала действия (А) и изометрического сокращения мышцы, приводящей большой палец кисти (Б).
1 — фаза развития напряжения; 2 — фаза расслабления.
152


 В
естественных условиях мышечные волокна
двигательной единицы
и скелетная мышца в целом работают в
режиме одиночного сокращения
только в том случае, когда длительность
интервала между
последовательными импульсами мотонейрона
равна или превышает
длительность одиночного сокращения
иннервируемых им мышечных
волокон. Так, режим одиночного сокращения
медленных волокон
камбаловидной мышцы человека обеспечивается
при частоте импульсации мотонейрона
менее 10 имп/с, а быстрых волокон
глазодвигательных
мышц — при частоте импульсации мотонейрона
менее
50 имп/с.
В
естественных условиях мышечные волокна
двигательной единицы
и скелетная мышца в целом работают в
режиме одиночного сокращения
только в том случае, когда длительность
интервала между
последовательными импульсами мотонейрона
равна или превышает
длительность одиночного сокращения
иннервируемых им мышечных
волокон. Так, режим одиночного сокращения
медленных волокон
камбаловидной мышцы человека обеспечивается
при частоте импульсации мотонейрона
менее 10 имп/с, а быстрых волокон
глазодвигательных
мышц — при частоте импульсации мотонейрона
менее
50 имп/с.
В режиме одиночного сокращения мышца способна работать длительное время без развития утомления. Однако в связи с тем, что длительность одиночного сокращения невелика, развиваемое мышечными волокнами напряжение не достигает максимально возможных величин. При относительно высокой частоте импульсации мотонейронов каждый последующий раздражающий импульс приходится на фазу предшествующего напряжения волокона, то есть до того момента, когда оно начинает расслабляться. В этом случае механические эффекты каждого предыдущего сокращения суммируются с последующим. Причем величина механического ответа на каждый последующий импульс меньше, чем на предыдущий. После нескольких первых импульсов последующие ответы мышечных волокон не изменяют достигнутого напряжения, а лишь поддерживают его. Такой режим сокращения называется гладким тетанусом (рис.4.6.). В подобном режиме двигательные единицы мышц человека работают при развитии максимальных изометрических усилий. При гладком тетанусе развиваемое ДЕ напряжение в 2-4 раза больше, чем при одиночных сокращениях.

Рис.4.6. Одиночные (а) и тетанические (б,в,г,д) сокращения скелетной мышцы. Накладывание волн сокращения друг на друга и образование тетануса при частотах раздражения: 5—15 раз в 1 с; в — 20 раз в 1 с; г — 25 раз в 1 с; д — более 40 раз в 1 с (гладкий тетанус).
153



 В
тех случаях, когда промежутки между
последовательными импульсами
мотонейрона меньше времени полного
цикла одиночного сокращения,
но больше длительности фазы напряжения,
сила сокращения
ДЕ колеблется. Этот режим сокращения
называется зубчатым
тетанусом (рис.
4.6.).
В
тех случаях, когда промежутки между
последовательными импульсами
мотонейрона меньше времени полного
цикла одиночного сокращения,
но больше длительности фазы напряжения,
сила сокращения
ДЕ колеблется. Этот режим сокращения
называется зубчатым
тетанусом (рис.
4.6.).
Гладкий тетанус для быстрых и медленных мыши достигается при разных частотах импульсации мотонейронов. Зависит это от времени одиночного сокращения. Так, гладкий тетанус для быстрой глазодвигательной мышцы проявляется при частотах свыше 150-200 имп/с, а у медленной камбаловидной мышцы — при частоте около 30 имп/с. В режиме тетанического сокращения мышца способна работать лишь короткое время. Это объясняется тем, что из-за отсутствия периода расслабления она не может восстановить свой энергетический потенциал и работает как бы "в долг".
Механическая реакция целой мышцы при ее возбуждении выражается в двух формах — в развитии напряжения и в укорочении. В естественных условиях деятельности в организме человека степень укорочения мышцы может быть различной. По величине укорочения различают три типа мышечного сокращения: 1. изотонический — это сокращение мышцы, при которой ее волокна укорачиваются при постоянной внешней нагрузке. В реальных движениях чисто изотоническое сокращение практически отсутствует; 2. изометрический — это тип активации мышцы, при котором она развивает напряжение без изменения своей длины. Изометрическое сокращение лежит в основе статической работы; 3. ауксотони-ч е с к и й или анизотонический тип — это режим, в котором мышца развивает напряжение и укорачивается. Именно такие сокращения имеют место в организме при естественных локомоциях — ходьбе, беге и т.д. Изотонический и анизотонический типы сокращения лежат в основе динамической работы локомоторного аппарата человека.
При динамической работе выделяют: 1. концентрический тип сокращения — когда внешняя нагрузка меньше, чем развиваемое мышцей напряжение. При этом она укорачивается и вызывает движение; 2. эксцентрический тип сокращения — когда внешняя нагрузка больше, чем напряжение мышцы. В этих условиях мышца, напрягаясь, все же растягивается (удлиняется), совершая при этом отрицательную (уступающую) динамическую работу.
Сила изометрического напряжения и длина мышцы. Покоящаяся мышца эластична и обладает упругостью. Следовательно, в определенных пределах, чем больше она растягивается, тем большее продольное напряжение в ней развивается. Изолированная мышца имеет равновесную длину, при которой ее упругое напряжение равно нулю. Зависимость между длиной мышцы и ее напряжением в покое называется кривой пассивного напряжения (рис.4.7.). Кривая напряжения нарастает тем круче, чем больше степень растяжения мышцы.
Степень предварительного растяжения определяет не только величину пассивного эластического напряжения покоящейся мышцы, но
154

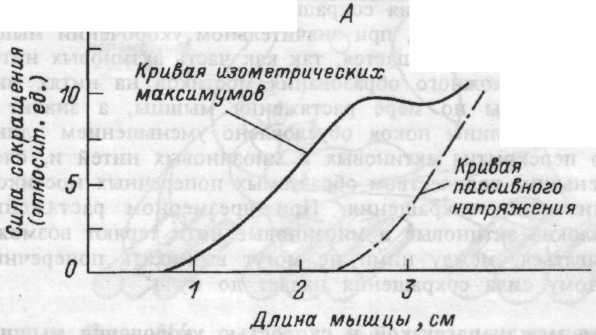
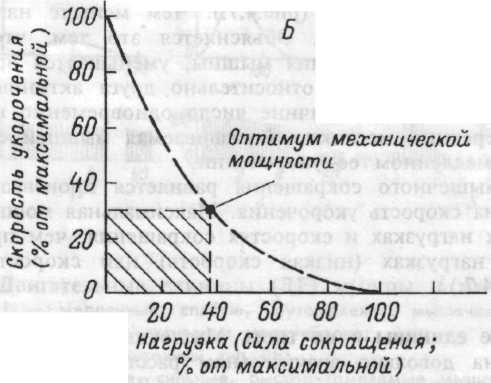
Рис.4.7. Зависимость между длиной мышцы и силой сокращения (А); между скоростью укорочения и величиной внешней нагрузки (Б).
и величину дополнительной силы, которую может развивать мышца в случае ее активации при данной исходной длине. Прирост силы при изометрическом сокращении суммируется с пассивным напряжением мышцы. Пиковые (максимальные) напряжения в этих условиях называют максимум изометрического напряжения (рис.4.7.). Напряжение сокращающейся мышцы максимально, если ее длина составляет примерно 120 % от равновесной. Это состояние носит название длины покоя.
Укорочение мышцы меньше длины покоя или ее растяжение больше этой величины приводит к снижению силы сокращения. Причина этого состоит в специфике взаимодействия актиновых и
155

 миозиновых
нитей. Поскольку напряжение, которое
развивают ми-офибриллы
в процессе развития сокращения, зависит
от числа поперечных
замкнутых мостиков, при значительном
укорочении мышцы
сила ее сокращения уменьшается, так как
часть актиновых нитей выходит
из зоны возможного образования мостиков
на нитях миозина.
Снижение силы по мере растяжения мышцы,
а значит и саркомеров,
больше длины покоя обусловлено уменьшением
длины зон
взаимного перекрытия актиновых и
миозиновых нитей и, следовательно,
меньшим количеством образуемых поперечных
мостиков, обеспечивающих
силу сокращения. При чрезмерном растяжении
мышечного
волокна актиновые и миозиновые нити
теряют возможность перекрываться,
между ними не могут возникать поперечные
мостики.
Поэтому сила сокращения падает до
нуля.
миозиновых
нитей. Поскольку напряжение, которое
развивают ми-офибриллы
в процессе развития сокращения, зависит
от числа поперечных
замкнутых мостиков, при значительном
укорочении мышцы
сила ее сокращения уменьшается, так как
часть актиновых нитей выходит
из зоны возможного образования мостиков
на нитях миозина.
Снижение силы по мере растяжения мышцы,
а значит и саркомеров,
больше длины покоя обусловлено уменьшением
длины зон
взаимного перекрытия актиновых и
миозиновых нитей и, следовательно,
меньшим количеством образуемых поперечных
мостиков, обеспечивающих
силу сокращения. При чрезмерном растяжении
мышечного
волокна актиновые и миозиновые нити
теряют возможность перекрываться,
между ними не могут возникать поперечные
мостики.
Поэтому сила сокращения падает до
нуля.