

5.2.2. Нециклический и циклический транспорт электронов
В фотосистеме IIреакционный центр, состоящий из димера хлорофиллаас максимумом поглощения при 680 нм (П680), последовательно поглотив энергию, равную 2 квантам коротковолнового красного света, и перейдя в синглетное возбужденное состояние, передает 2 электрона феофитину. От феофитина электроны последовательно передаются на пластохиноныQAиQB, расположенные на другой стороне мембраны тилакоида, затем на две липидорастворимые молекулы пластохинонаPQ. Он является переносчиком протонов и электронов, выполняющего в мембране роль челнока. Принимая 2 электрона и захватывая из стромы 2 Н+, 2 молекулыPQH2диффундируют через липидную фазу мембраны. Электроны от 2PQH2поступают в цепь:
железосерный белок FeSrцитохромfCu-содержащий белок пластоцианинП700,
а протоны попадают в полость тилакоида.
Вакантные места в П+680заполняются 2 электронами из содержащегоMnпереносчика электронов, который восстанавливается с участием белкового комплексаS. Окисленный комплексSсвязывает воду и восстанавливается за счет электронов воды. При фотоокислении воды комплексомSвторая пара протонов освобождается в полость тилакоида, а атом кислорода диффундирует из хлоропласта.
После возбуждения реакционного центра фотосистемы I- молекулы хлорофиллаа с максимумом поглощения при 700 нм (П700), 2 квантами длинноволного красного света 2 электрона передаются мономерной форме хлорофиллаа – А1и затем переносчикам электронoв железосерным белкам А2и АВ, ферредоксину на наружной стороне мембраны тилакоида, НАДФ-редуктазе. Этот фермент восстанавливает НАДФ в НАДФН. На вакантные места в П+700переходят электроны с пластоцианина и нециклическая цепь транспорта электронов замыкается.
В мембранах хлоропластов может функционировать циклический транспорт электронов, когда действует только фотосистема I. Возбужденные молекулы П700передают электроны на А1, железосерные белки А2и АВ, переносящие электроны на другую сторону мембраны, ферредоксин, пластохинон, цитохромb6, железосерный белокFeSr, цитохромf, пластоцианин и П700(рис. 5.4). Энергия, освобождающаяся при транспорте электронов, используется для фосфорилирования АДФ.
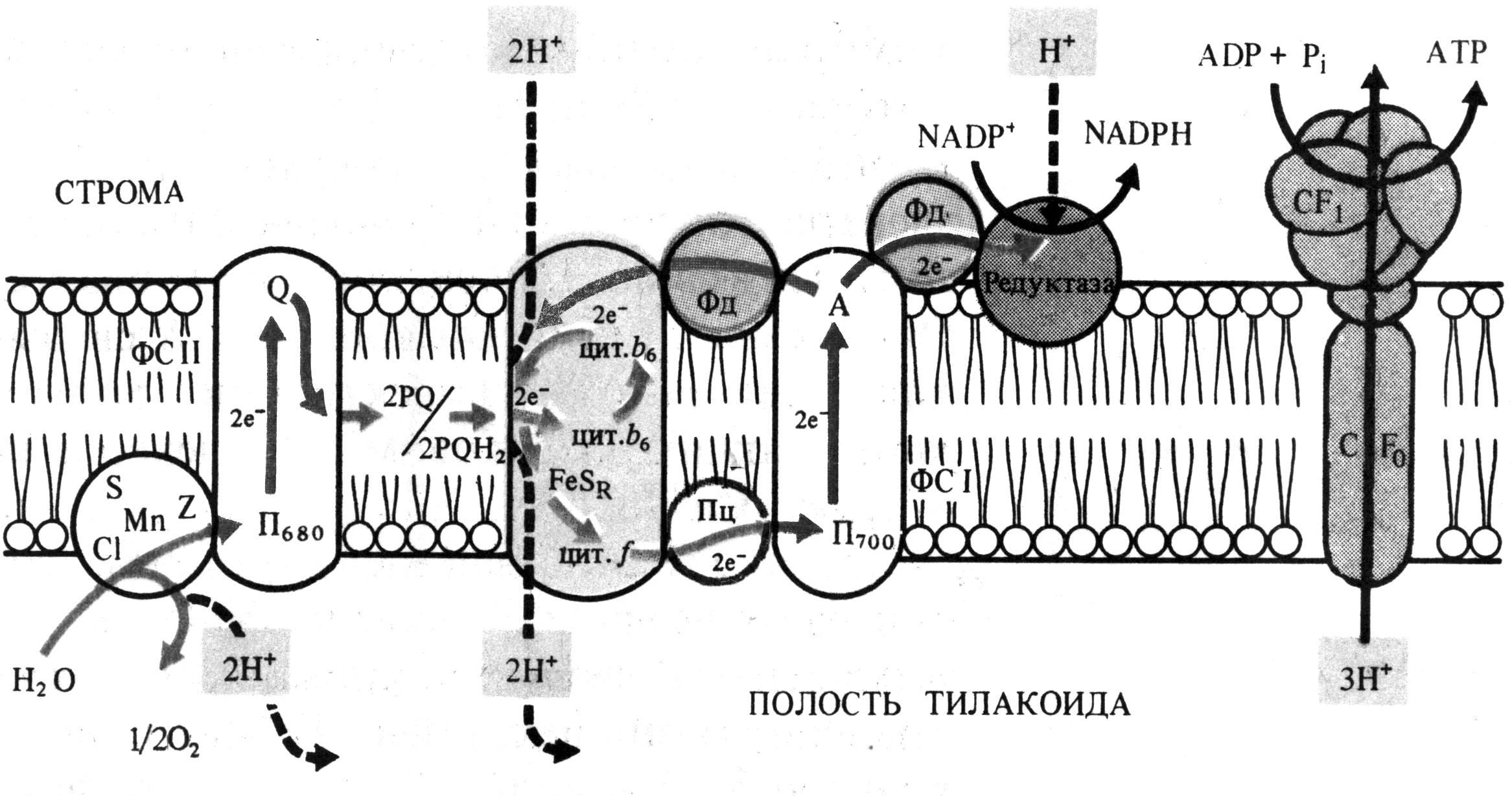
Рис. 5.4. Локализация электрон- и протонтранспортных реакций в тилакоидной мембране хлоропласта (по В. В. Полевому).
Механизм фотофосфорилирования АДФ объясняет теория П. Митчелла. Согласно этой теории, трансмембранный перенос пластохинонами электронов и протонов в одну сторону чередуется с переносом цитохромной системой в обратную сторону электронов. Поэтому по одну сторону мембраны накапливается избыток протонов и возникает электрохимический мембранный потенциал. Его энергия используется для синтеза АТФ при разрядке мембраны в результате транспорта протонов через мембрану посредством Н+-АТФазы, которая действует как АТФсинтетаза. Этот фермент выглядит как грибовидная частица на поверхности мембраны тилакоидов. Его молекула состоит из двух частей: головки, выступающей с наружной стороны мембраны и содержащей активный центр фермента, и ножки, погруженной в мембрану и представляющей канал, через который передвигаются протоны.