

69_39_Genetikapdf
.pdf31
У метафазі I в екваторіальній площині клітини чітко видні біваленти, прикріплені центромерами до ниток веретена поділу.
Рис. 2.8. Схема мейозу
В анафазі мейозу I гомологічні хромосоми, що складаються з двох хроматид, відходять до протилежних полюсів клітини. Розбіжність хромосом носить випадковий характер.
Телофаза мейозу I не відрізняється від такої мітозу (але хромосоми не деспіралізуються).
У результаті мейозуI утворюються дві дочірні клітини, що містять гаплоїдний набір хромосом, але кожна хромосома має дві хроматиди. Отже, у результаті мейозу I відбувається редукція (зменшення вдвічі) числа хромосом, звідки і назва цього поділу - редукційне.
Після закінчення мейозу I настає короткий проміжок - інтеркінез, протягом якого не відбуваються реплікація ДНК і подвоєння хроматид.
32
Мейоз II протікає по типу звичайного мітозу. Профаза мейозу II нетривала,
тому що |
хромосоми |
після телофази мейозуI залишаються спіралізованими. |
У |
метафазі |
мейозу II |
хромосоми розташовуються в екваторіальній площині |
|
клітини. У анафазі мейозу II відбувається поділ хроматид в області центромер і |
|||
до полюсів відходять хроматиди (дочірні хромосоми) У телофазі мейозу II після |
|||
цитокінеза утворюються клітини з гаплоїдним набором хромосом. |
|
||
Таким чином, у результаті двох послідовних поділів мейозу |
з однієї |
||
диплоїдної клітини утворяться 4 гаплоїдні. Значення мейозу: 1) редукція числа |
|||
хромосом, |
2) кон’югація гомологічних хромосом, 3) рекомбінація генетичного |
||
матеріалу, обумовлена кросинговером і випадкавою розходженням гомологічних |
|||
хромосом. |
Отже, завдяки мейозу забезпечується підтримка сталості |
числа |
|
хромосом при наявності статевого процесу, а також комбінативна мінливість. |
|
||
Запитання для самоперевірки:
1.Які положення включає сучасна клітинна теорія?
2.Що таке цитоплазматична мембрана і яка її роль у житті клітини? Які властивості та функції мембран?
3.Як відбувається пасивний транспорт речовин у клітину? В чому суть активного транспорту речовин у клітину?
4.Яким чином використовуються речовини, що надійшли у клітину?
5.Які органели клітини відносяться до анаболічної системи?
6.Що входить до катаболічної системи клітини?
7.У чому відмінність еукаріотичної клітини від прокаріотичної?
8.Охарактеризуйте структуру клітинного ядра.
9.Що вивчає цитогенетика? Які ви знаєте сучасні методи цитогенетичних досліджень?
10.Що таке хромосома? Які основні функції хромосом?
11.Охарактеризуйте метафазну хромосому.
12.Які бувають типи хромосом у залежності від розташування центромери?
13.Як поділяють хромосоми за стадіями мітозу? Що таке гетерохромосоми та аутосоми?
14.Поясніть поняття «гаплоїндний» і «диплоїдний» набір хромосом.
15. Охарактеризуйте |
хромосоми |
типу |
лампових |
щіток |
і |
політ |
хромосоми. |
|
|
|
|
|
|
16.Охарактеризуйте перший рівень упакування ДНК.
17.Який рівень упаковки ДНК складає основу хроматиди?
18.Що таке клітинний та мітотичний цикли?
19.Який основний спосіб поділу соматичних клітин?
20.Які головні причини початку мітозу? Скільки і які стадії мітозу відомі? У чому полягає значення мітозу? Чи є мітоз єдиним способом поділу клітин?
21.Що таке мейоз і які його етапи? Що таке кросинговер і на якому етапі мейозу він відбувається? Чому мейоз І називають редукційним поділом?
22.Яке значення мейозу?

33
ТЕМА 3. ХРОМОСОМНА ТЕОРІЯ СПАДКОВОСТІ
3.1. Хромосоми як групи щеплення генів
У. |
Сеттон |
і |
. РПеннет у 1908 |
р. знайшли |
відхилення |
від |
вільного |
|
|||||
комбінування ознак відповідно до третього закону Менделя. У 1911-1912 рр. Т. |
|
|
|||||||||||
Морган зі співробітниками описали явище зчеплення генів- спільну передачу |
|
||||||||||||
групи генів з покоління в покоління. Досліди проводилися на мухах дрозофілах з |
|
||||||||||||
обліком двох пар альтернативних ознак- сірий і чорний колір тіла, нормальні і |
|
|
|||||||||||
короткі |
крила. |
При |
схрещуванні |
|
гомозиготних |
особин |
із |
сірим |
тілом |
|
|||
нормальними крилами з особинами з чорним |
тілом і |
короткими |
крила |
||||||||||
отримана |
однаковість |
гібридів |
першого |
покоління, особини |
якого |
мали |
|
||||||
домінантні ознаки. |
|
|
|
|
|
|
|
|
|
|
|
||
Для з’ясування генотипу гібридівI покоління Морган провів аналізуюче |
|
||||||||||||
схрещування. Він |
узяв |
рецесивну |
гомозиготну |
самицю |
і |
схрестив |
ї |
||||||
дигетерозиготним самцем, потім провів реципрокне схрещування. При вільному комбінуванні генів відповідно до третього закону Менделя в поколінні повинні були з’явитися мухи чотирьох різних фенотипів по25%. Це можна пояснити наступною схемою:
Але гени різних |
алельних |
пар |
можуть |
знаходитися |
у одній |
|
гомологічних |
хромосом. |
Одна з |
пари |
гомологічних хромосом |
містить2 |
|
домінантних |
гена (BV), а |
інша - 2 рецесивних (bv). У |
процесі мейозу |
одна |
||
хромосома з генами ВV потрапить в одну гамету, а інша (з генами bv) - в іншу. Таким чином, у дигетерозиготного організму утвориться не чотири, а тільки два типи гамет і нащадки будуть мати таке ж сполучення ознак, як і батьки. У даному
випадку зчеплення буде повним. |
|
При вивченні результатів |
другого схрещування(II) було виявлене |
порушення повного зчеплення генів. |
Якщо взяти дигетерозиготну самицю мухи |
дрозофіли і схрестити її з рецесивним самцем, то виходить 4 різновиди фенотипів нащадків: 41,5% із сірим тілом і довгими крилами, 41,5% з чорним тілом і короткими крилами і по8,5% гібридних форм - із сірим тілом і короткими крилами і з чорним тілом і довгими крилами. У цьому випадку зчеплення виявляється неповним, тобто відбувається перекомбінація генів, локалізованих в одній хромосомі. Це пояснюється кросинговером гомологічних хромосом.
Гени, |
локалізовані в |
одній хромосомі, передаються разом (зчеплено) і |
||||
складають |
одну |
групу |
зчеплення. |
Оскільки |
в |
гомологічних хромосомах |
локалізовані алельні гени, групу зчеплення складають дві гомологічні хромосоми |
||||||
і кількість |
груп |
зчеплення |
дорівнює |
кількості |
пар |
хромосом(чи гаплоїдному |
34
числу хромосом). Так, у мухи дрозофіли8 хромосом - 4 групи зчеплення, у людини 46 хромосом - 23 групи зчеплення.
3.2. Кросинговер
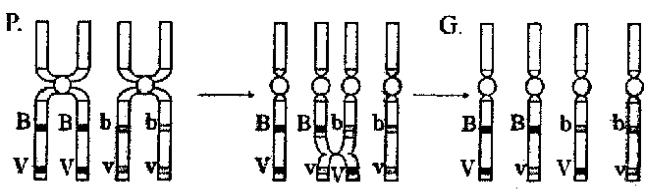
Кросинговер - обміном ділянками гомологічних хромосом у процесі їхньої кон’югації у профазі мейозу I (рис. 3.1). Кожна з хроматид, що утворилися, потрапляє в окрему гамету. Утворюється чотири типи гамет, але на відміну від вільного комбінування їх процентне співвідношення не буде рівним, тому що кросинговер відбувається не завжди.
Рис. 3.1. Схема кросинговеру
Сила зчеплення між генами(частота кросинговера) залежить від відстані між ними: чим більше відстань, тим менше сили зчеплення, тим частіше відбувається кросинговер. Чим більшою є відстань між двома хромосомними локусами, тим більше рекомбінаційних подій у більшій кількості точок може відбутися між ними. Це означає, що частота кросинговеру є мірою фізичної відстані між локусами, і у відсотках кросинговеру можна вимірювати відносну відстань між генами на хромосомі.
Відстань між генами визначається за відсотком кросинговера. За одиницю
відстані приймається одна |
|
морганіда(на |
честь |
Моргана), |
що дорівнює 1% |
|||||||||
кросинговера. Гамети, у які потрапили хроматиди, але не пройшли кросинговер, |
||||||||||||||
називаються |
некросоверними, |
їх |
звичайно |
більше. Гамети, у |
які |
потрапили |
||||||||
хроматиди після кросинговеру, називаються кросоверними, їх звичайно менше. |
||||||||||||||
Кросинговер унаслідок |
гомологічної |
рекомбінації |
відбувається |
тільки в |
||||||||||
парах гомологічних хромосом, які є однаковими за морфологією та набором генів |
||||||||||||||
у обох статей, – так званих аутосом. У більшості організмів, які розмножуються |
||||||||||||||
статевим |
шляхом, |
одна |
|
|
пара |
хромосом(статеві) |
представлена |
|
двома |
|||||
негомологічними |
типами. |
Одна |
зі |
статей |
|
при |
(цьомугомогаметна) |
|||||||
характеризується |
двома |
|
|
однаковими |
|
статевими |
хромосомами, інша |
|||||||
(гетерогаметна) – двома |
різними (наприклад, |
у |
ссавців |
двіX-хромосоми |
||||||||||
визначають |
самку, |
X- |
і |
Y-хромосома |
– самця). |
Рекомбінація |
в |
парі |
||||||
негомологічних статевих хромосом є неможливою (або суттєво обмеженою), отже статеві хромосоми передаються як одне ціле від гетерогаметної статі до нащадка.
35
Власне, у перемішуванні генів між двома гомологічними хромосомами і полягає основна біологічна роль кросинговеру. Кросинговер забезпечує для
багатьох пар генів практично незалежне |
спадкування. З огляду |
на , тещо |
|
||||
більшість рецесивних алелів є мутантними, кросинговер різко підвищує частку |
|
||||||
життєздатних |
гетерозигот |
у |
наступному |
поколінні. Крім того, |
викликане |
|
|
кросинговером |
незалежне |
спадкування |
генів зумовлює |
створення |
велик |
||
кількості їхніх комбінацій у межах популяції, що сприяє, з одного боку, загальній |
|
||||||
генетичній стабільності популяції, |
з іншого – |
забезпечує певний |
еволюційний |
|
|||
потенціал. |
|
|
|
|
|
|
|
Кросинговер відбувається в усіх досліджених видів тварин і рослин, хоча наявність процесу та його частота може залежати від статі. Частота кросинговеру залежить також від факторів зовнішнього середовища і стадій індивідуального розвитку. Уздовж хромосоми частоти гомологічної рекомбінації розподілені нерівномірно: залежність частоти від фізичної відстані є лінійною в середніх частинах плечей хромосом, ближче до гетерохроматинових центромерних і теломерних зон кросинговер пригнічується. Існують також певні «гарячі точки», які характеризуються підвищеною частотою рекомбінації. Нарешті, як і будь-який інший біологічний процес, гомологічна рекомбінація контролюється певним набором генів, мутації яких також впливають на частоту кросинговеру.
Гомологічна рекомбінація – досить точний процес, який відбувається між гомологічними ділянками ДНК, у результаті відбувається обмін однаковими за довжиною ділянками, що містять однакову кількість . генівАле іноді спостерігається рекомбінація між гомологічними ділянками, які займають різне положення на хромосомі, унаслідок чого здійснюється обмін неоднаковими за довжиною ділянками – нерівний кросинговер. Такий обмін зумовлює дуплікацію ділянки в одній хромосомі та делецію в іншій. Таким чином, нерівний кросинговер є однією з причин хромосомних перебудов.
3.3. Основні положення хромосомної теорії спадковості
Основні положення хромосомної теорії спадковості(Т. Морган і співавт., 1911 р.) зводяться до наступного:
·Гени розташовані в хромосомах у лінійному порядку у певних локусах. Алельні гени займають однакові локуси гомологічних хромосом.
·Гени, розташовані в одній хромосомі, утворюють групу зчеплення й успадковуються переважно разом; число груп зчеплення дорівнює гаплоїдному набору хромосом.
·Між гомологічними хромосомами можливий обмін ділянками- кросинговер, що порушує зчеплення генів.
·Відсоток кросинговера пропорційний відстані між генами. 1 морганіда - одиниця відстані, рівна 1% кросинговера.

36
3.4. Картування хромосом: цитологічна та генетична карта хромосоми
Знаючи відстань між генами можна побудувати карту хромосо. Генетична карта хромосоми – це відрізок прямої, на якому позначений порядок розташування генів і зазначена відстань між ними у морганідах. Вона будується за результатами аналізуючого схрещування.
Цитологічна карта хромосоми– це фотографія чи точний малюнок хромосоми, на якому відзначається послідовність розташування генів. Її будують на основі зіставлення результатів аналізуючого схрещування, і хромосомних перебудов (рис. 3.2). Наприклад, якщо хромосома з домінантними генами буде послідовно втрачати окремі локуси(при впливі на неї мутагенів), то в гетерозиготі почнуть виявлятися рецесивні ознаки, порядок появи ознак буде вказувати на послідовність розташування генів. Цитологічні карти створюються шляхом визначення локалізації генів у хромосомах.
Рис. 3.2. Схема генетичної (А) і цитологічної (В) карт хромосом
Запитання для самоперевірки:
1.Що таке група зчеплення генів? Від чого залежить сила зчеплення між генами?
2.Чому кількість груп зчеплення дорівнює кількості пар хромосом?
3.Що таке кросинговер? Між якими хромосомами можливий кросинговер?
4.Чи можливий кросинговер у парі негомологічних статевих хромосом?
5.У яких одиницях визначається відстань між генами?
6.У чому полягає основна біологічна роль кросинговеру?
7.Від яких факторів залежить частота кросинговеру?
8.Що таке нерівний кросинговер? Які його наслідки?
9.Назвіть основні положення хромосомної теорії спадковості.
10.Що показує генетична карта хромосоми?
11.Як створюються цитологічні карти хромосом?
37
ТЕМА 4. ПОЗАЯДЕРНА СПАДКОВІСТЬ
4.1. Геном мітохондрій і хлоропластів
Переважна більшість еукаріотичних генів міститься в клітинному ядрі. Крім ядра, свій власний генетичний матеріал мають цитоплазматичні органели еукаріотичної клітини: мітохондрії та хлоропласти.
За своєю організацією геноми органел значно відрізняються від ядерного геному й нагадують геноми прокаріотів. Це стало основоюендосимбіотичної теорії походження мітохондрій і пластид, відповідно до якої вільно існуючі -α протобактерії та ціанобактерії були поглинуті протоеукаріотичною клітиною і еволюціонували у специфічні органели(мітохондрії та хлоропласти), які відповідають за дихання та фотосинтез.
Мітохондріальний геном може бути представлений однією або кількома молекулами ДНК, які в комплексі з білками утворюють структуру, подібну до нуклеоїдів прокаріотів. Мітохондріальна ДНК може бути як кільцевою, так і лінійною, причому лінійні мтДНК характеризуються наявністю специфічних теломероподібних кінцевих структур.
Мітохондріальні геноми значно варіюють за розмірами: від 6 тис. пар основ у плазмодіїв до 2,5 млн пар основ у дині. Різниця в розмірах мтДНК викликана
наявністю некодуючих послідовностей, що є особливо характерним для вищих рослин (50 – 70 %). Також характерним для мтДНК рослин є наявність вбудованих ділянок хлоропластної ДНК. У тварин мтДНК має в основному невеликі розміри (13 – 19 тис. пар основ).
Набір генів у мтДНК варіює від 5 до 100. Для більшості еукаріотів середня
кількість генів мтДНК становить40 – 50, з |
яких 12 – 20 є білковими. Геноми |
||||||||
мітохондрій усіх еукаріотів кодують рРНК великої та |
малої |
субодин |
|||||||
мітохондріальної рибосоми, а також частковий(інколи повний) набір власних |
|||||||||
тРНК. Білки, |
що |
кодуються |
мтДНК, в |
основному |
залучені |
до |
процесів |
||
перенесення електронів та синтезу АТФ. Генетичний код, що використовується |
|||||||||
власною |
системою |
трансляції |
мітохондрій, характеризується |
деякими |
|||||
відхиленнями від універсальної таблиці відповідності кодонів амінокислотам. |
|
||||||||
Усі |
компоненти |
|
реплікативного, транскрипційного |
та |
частина |
||||
трансляційного апаратів |
мітохондрій |
кодуються |
ядерним |
геномом. Отже, |
|||||
експресія мітохондріальних генів перебуває під повним контролем ядра. |
|
||||||||
Хлоропласти |
вищих |
рослин містять |
багато |
ідентичних |
кільцевих |
||||
дволанцюгових молекул ДНК, розміри яких коливаються від120 до 220 тис. пар основ. Характерною особливістю хлоропластної ДНК(хлДНК) вищих рослин є наявність інвертованого повтору (ІП), довжина якого варіює в різних видів від5 до 76 тис. пар основ. Відмінності за розмірами хлДНК різних видів в основному визначаються довжиною ІП. Як правило, ІП представлений двома сегментами (ІП- 1 та ІП-2), що розділяють хлДНК на велику й малу унікальні ділянки. Велика унікальна ділянка є найбільш варіабельною частиною молекули хлДНК.
38
За своєю організацією хлоропластний геном подібний до бактеріального. Основні регуляторні послідовності, такі як промотори і термінатори, в обох геномах фактично ідентичні. Разом із тим, на відміну від прокаріотів, у генах хлДНК присутні інтрони.
За складом і порядком розміщення генів хлоропластні геноми вищих рослин є високо консервативними. У вивчених геномах хлоропластів вищих рослин
виявлено від 108 до 122 генів, 95 з яких є однаковими й зустрічаються в усіх видів.
За функціональністю гени хлоропластів можна розділити на три групи:
·гени апаратів транскрипції та трансляції;
·гени, що пов’язані з фотосинтезом;
·гени фотосинтетичного метаболізму(біосинтезу амінокислот, жирних кислот, пігментів тощо).
Усі найважливіші білки, які беруть участь у реплікації, транскрипції та трансляції кодуються ядерним геномом. Тобто, як і для мітохондрій, усі процеси,
що відбуваються у хлоропластах, перебувають під контролем ядра.
4.2. Пластидна, мітохондріальна та псевдоцитоплазматична спадковість
Цитоплазматична |
спадковість – |
це |
спадковість, |
яка контролюється в |
основному генами, що |
розміщені |
у |
неядерних |
органелах(мітохондріях, |
хлоропластах і т. д.)
Поряд з ядерними генами, локалізованими в хромосомах, виявлені фактори спадковості, розташовані у цитоплазмі. Їх називають плазмогенами (плазмідами). Хімічну основу плазмогенів складають молекули . ДНКУстановлено, що пластиди, мітохондрії і деякі інші органоїди містять ДНК. У цитоплазмі може знаходитися і чужорідна ДНК вірусів і плазміди бактерій. Позаядерна ДНК здатна реплікуватися незалежно від реплікації хромосом. Плазмогени знаходяться під контролем ядерних генів. Цитоплазматичне успадкування іде по материнській лінії, тобто через цитоплазму яйцеклітини, тому що сперматозоїд майже її не містить. Критеріями цитоплазматичної спадковості є:
·відсутність кількісного менделевського розщеплення в потомстві;
·неможливість виявити зчеплення;
·різні результати реципрокних схрещувань.
Виділяють наступні основні види цитоплазматичної спадковості: пластидну, мітохондріальну і псевдоцитоплазматичну.
Відкриття пластидної спадковості належить К. Корренсу (1908), що описав строкатість у нічної красуні. У строкатих рослин частина пластид нездатна
утворювати |
хлорофіл. Пластиди |
при |
мітозі розподіляються між |
дочірніми |
|
клітинами |
нерівномірно. |
Частина |
клітин одержує тільки нормальні пластиди |
||
(листя будуть зеленими); |
частина |
клітин |
одержує тільки аномальні |
пластиди |
|
(листя білі, без хлорофілу, рослина гине); нарешті клітини дістають й аномальні і нормальні пластиди (строкате листя, білі плями на зелених листах).
39
Мітохондріальна спадковість описана Б. Ефруссі (1949). Він знайшов, що близько 1% хлібних дріжджів дають карликові колонії. Виявилося, що клітини карликових колоній не мають у мітохондріях дихальних ферментів внаслідок мутації плазмогенів і тому ростуть дуже повільно. Гени, що кодують дихальні ферменти, знаходяться в кільцевих молекулах ДНК мітохондрії. Довжина кожної такої молекули приблизно 15000 пар нуклеотидів. Розрахунки показали, що обсяг
власної спадкової |
інформації мітохондрії недостатній для відтворення всієї |
сукупності РНК і |
білків органоїда. Багато білків, запрограмованих ядерними |
генами, включаються в структуру мітохондрії.
Геном мітохондрії людини представлений кільцевою молекулою ДНК, що містить 16569 пар нуклеотидів. До складу генома входять також гени рРНК, 22 різних т-РНК, субодиниць I, II і III оксидази цитохрому С, субодиниці 6 АТФази, цитохрома b і дев’яти інших поки що невідомих білків. У мітохондріальній ДНК є дуже мало некодуюючих ділянок і обидва її ланцюжки транскрибуються. Є дані про те, що деякі вади розвитку людини обумовлені мутаціями мітохондріальних генів (мітохондріальна цитопатія, незрощення верхніх дуг хребців і зрощення нижніх кінцівок, паркінсонізм і ін.).
У цитоплазмі бактерій виявлені автономно розташованіплазміди, що складаються з кільцевих молекул дволанцюгової .ДНКВони обумовлюють стійкість бактерій до ліків(антибіотиків), програмують синтез деяких отрут (гемолізину, ентеротоксину). Плазміди забезпечують також обмін генетичною інформацією між мікроорганізмами. Позахромосомні молекули ДНК широко
використовуються в генній інженерії, тому що вони здатні містити в |
собі |
||
генетичний матеріал хромосом і переносити його в інші клітини. |
|
|
|
Псевдоцитоплазматична |
спадковість обумовлена |
проникненням |
у |
цитоплазму клітин ділянок чужорідної ДНК, тобто вона являє собою різновид
внутрішньоклітинного |
паразитизму. Так, у деяких |
ліній |
мух дрозофіл існує |
підвищена чутливість |
до 2СО. Установлено, що |
ця |
здатність обумовлена |
передачею особливих вірусів через цитоплазму яйцеклітини. |
|
||
Умишей описані лінії зі спадковою схильністю до розвитку раку молочної залози. При детальному вивченні цього явища встановлено, щ схильність передається не через статеві клітини, а через молоко, у якому міститься вірус (фактор молока). Якщо новонароджених мишенят «ракової» лінії вигодовує самка «нормальної» лінії, вони залишаються здоровими. Якщо мишеняти «нормальної» лінії вигодовує самка «ракової», то в останніх розвивається рак молочної залози.
4.3.Основні характеристики спадкування генів органел
Удиплоїдних видів еукаріотів ядерні гени зумовлюють успадкування ознак за законами Менделя в результаті того, що, за винятком зчеплених зі статтю
ознак, нащадки отримують по одній копії |
кожного гена |
від обох . батьк |
Мітохондрії та пластиди не розподіляються |
по гаметах і |
дочірніх клітина |
аналогічно хромосомам ядра, тому закономірності спадкування генів органел і ознак, які вони зумовлюють, значно відхиляються від менделівських. Оскільки
40
мітохондрії та пластиди знаходяться в цитоплазмі клітин, такий тип спадкування називається позаядерним (позахромосомним), або цитоплазматичним.
Особливості позаядерного спадкування зумовлені в першу чергу тим, що
гамети різних статей мають непорівнянні кількості мітохондрій і |
. пласти |
Найчастіше жіночі гамети містять тисячі мітохондрій( пластид у рослин), |
а |
чоловічі – одиниці. У результаті в зиготі переважають мітохондрії та хлоропласти |
|||||||||
матері. У деяких видів, зокрема в людини, чоловічі мітохондрії взагалі не |
|||||||||
потрапляють |
до |
зиготи. Отже, спадкування |
|
генів |
органел |
відбувається в |
|||
основному по материнській лінії. Проте це |
загальне |
правило |
не носить |
||||||
абсолютного |
характеру – для деяких |
видів |
гризунів |
спостерігається |
передача |
||||
мітохондрій у зиготу від обох батьків. |
|
|
|
|
|
|
|
||
Особливості спадкування хлоропластних генів були вперше вивчені при |
|||||||||
дослідженні спадкування строкатості листків нічної красуні. Якщо в ролі |
|||||||||
материнської форми брали квіти безхлорофільної рослини і запилювали їх пилком |
|||||||||
зеленої рослини, то |
у F1 спостерігали |
тільки |
безхлорофільні |
форми(незабаром |
|||||
гинуть). При реципрокному схрещуванні уF1 усі рослини нормальні. Якщо |
|||||||||
запилювали квіти строкатих рослин пилком зеленої форми, у F1 виявляли |
|||||||||
безхлорофільні, строкаті й зелені рослини. При |
реципрокному |
схрещуванні– |
|||||||
тільки зелені. |
|
|
|
|
|
|
Таким |
чином, розглянутий |
приклад |
демонструє |
материнський |
тип |
|
спадкування |
хлоропластного геному в рослин, для якого |
є характерною |
||||
контрастна різниця між результатами реципрокних схрещувань. |
|
|
||||
Досить схожі відмінності в таких схрещуваннях спостерігаються при |
||||||
схрещуванні зелених і строкатих форм у інших рослин. Проте все залежить від |
|
|||||
кількостей |
цитоплазми, що |
привносяться |
в |
зиготу |
яйцеклітиною |
|
сперматозоїдом. Так, якщо квіти строкатих рослин герані запилюють пилком зеленої, то до 30 % нащадків будуть строкатими, а 70 % – зеленими. При
реципрокному схрещуванні 70 % нащадків є |
строкатими, а 30 % – зеленими. |
Тобто в рослин спостерігається також |
і двобатьківське спадкування ген |
хлоропластів. |
|
Запитання для самоперевірки:
1.Які органели еукаріотичної клітини містять генетичний матеріал?
2.Охарактеризуйте мітохондріальній геном.
3.Поясніть чому експресія мітохондріальних генів перебуває під повним контролем ядра.
4.Які ви знаєте особливості хлоропласної ДНК? Охарактеризуйте гени хлоропластів.
5.Що таке цитоплазматична спадковість?
6.Які критерії цитоплазматичної спадковості?
7.Поясніть явище пластидної спадковості.
8.Чим характеризується мітохондріальна спадковість?
9.Чим обумовлена псевдоцитоплазматична спадковість?
10.Чим зумовлені особливості позаядерного спадкування генів?