

69_39_Genetikapdf
.pdf61
ми термінації трансляції– жодна тРНК не містить відповідних антикодонів. Фактори термінації забезпечують звільнення синтезованого амінокислотного ланцюга та дисоціацію рибосоми.
7.3. Відмінності між іРНК, а також процесів транскрипції і трансляції у прота еукаріот
Суттєвою особливістю прокаріотичної системи транскрипції/трансляції є те, що молекула іРНК зв’язується з рибосомами безпосередньо під час транскрипції –
прокаріотична транскрипція |
і білковий синтез(трансляція) є єдиним |
процесом. Прокаріотична іРНК |
єполіцистронною – містить інформацію про |
синтез кількох білків. Вона має кілька послідовних рамок зчитування та кілька стартових кодонів.
Специфічне розміщення стартового кодона в Р-сайті рибосоми при ініціації трансляції у прокаріотів забезпечується комплементарним упізнаванням між
ділянкою |
16S |
рРНК і |
консервативноюпослідовністю Шайна-Дальгарно, |
||
розміщеною в іРНК за 5 – 9 нуклеотидів від стартового кодона в напрямку до5´- |
|||||
кінця. Саме |
наявність |
цієї послідовності й робить кодон АУГ стартови, |
|||
відрізняючи його від звичайного метионінового кодона. Якщо рибосоми з тих чи |
|||||
інших причин не зв’язуються з іРНК, транскрипт |
швидко деградує |
під дією |
|||
нуклеаз. |
|
|
|
|
|
На |
відміну від |
прокаріотів, еукаріотична |
іРНК синтезується |
під час |
|
транскрипції у клітинному ядрі, звідки транспортується до цитоплазми. Отже, у
еукаріот білковий синтез (трансляція), який відбувається в цитоплазмі, та
транскрипція є окремими, розділеними у просторі й часі, етапами експресії
гена. |
|
|
моноцистронна (містить |
|
|
||
Еукаріотична |
іРНК – |
інформацію |
про |
||||
амінокислотну послідовність однієї білкової молекули). Кожна еукаріотична |
|||||||
молекула іРНК має тільки одну рамку зчитування(один стартовий кодон). |
|||||||
Упізнавання |
стартового |
кодона |
залежить |
від |
контексту |
нуклео |
|
послідовності, в якій він розташований (при цьому не обов’язково перший кодон |
|||||||
АУГ, що зустрічається, сприймається як стартовий). Найкращим контекстом, який |
|||||||
максимально |
|
сприяє |
ініціації |
трансляції, є послідовність |
Козак: |
||
ГЦЦ(А/Г)ЦЦАУГГ. |
|
|
|
|
|
|
|
Інша принципова відмінність полягає в мозаїчності будови еукаріотичного |
|
||||||
гену: наявність |
кодуючих |
ділянок– екзонів |
і некодуючих– інтронів. Під час |
|
|||
транскрипції |
спочатку |
синтезується |
|
пре-іРНК. Перетворення |
пре-іРНК |
у |
|
функціональну |
іРНК |
називають |
процесингом. Усі |
операції |
процесингу |
||
відбуваються |
під час |
транскрипції |
на |
РНК-полімеразному |
комплексі, тобто |
|
|
процесинг еукаріотичної іРНК є невід’ємною частиною транскрипції(рис. 7.5). Процесинг полягає в ферментативному розрізанні первинного транскрипту
про-іРНК з подальшим |
видаленням його |
інтронних |
ділянок і |
поєднанн |
(сплайсингу) екзонних ділянок, що формують безперервну кодуючу послідовність |
||||
зрілої іРНК, яка надалі бере |
участь у трансляції |
генетичної |
інформації. Під |
час |
62
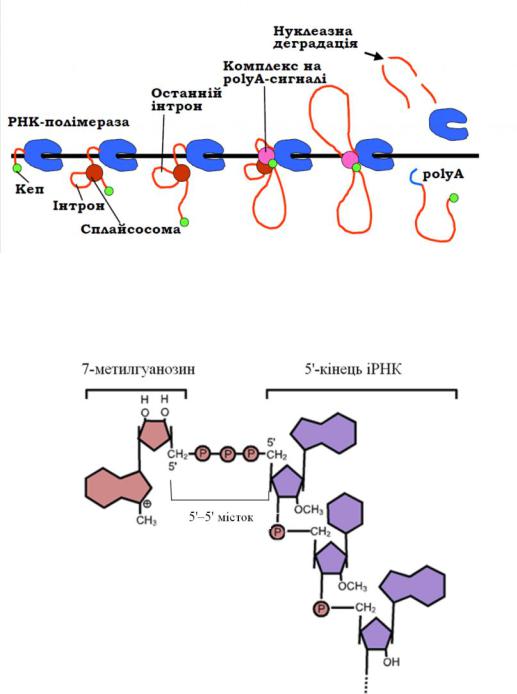
процесингу відбувається також модифікація5'- і 3'-кінців зрілої молекули іРНК. Модифікація 5'-кінця іРНК полягає в утворенні 5'-кінцевого кепу.
Рис. 7.5. Синхронізація транскрипції та процесингу іРНК
Кепування – це приєднання до5'-кінця транс-крипту РНК7-метил- гуанозину через незвичайний для РНК5´,5´-трифосфатних місток, а також метилювання залишків рибози двох перших нуклеотидів (рис. 7.6).
Рис. 7.6. Схема будови кепу іРНК
Після синтезу перших20 – 30 нуклеотидів пре-іРНК відбувається модифікація її 5´-кінцевого нуклеотиду з утворенням кепу (від англ. cap – шапка). Функціональне значення кепу: стимулює інші реакції процесингу, захищає іРНК від нуклеазної деградації, приймає участь у транспорті іРНК з ядра до цитоплазми, бере участь у ініціації трансляції. Далі під час елонгації транскрипції, після синтезу чергового інтрону, відбувається збирання мультимолекулярної структури, яка називається сплайсосомою. Призначення сплайсосоми полягає у здійсненні сплайсингу – вирізання інтронів і зшивання екзонів, у результаті чого

63
іРНК стає копією лише кодуючої частини гену або її фрагментів: сплайсинг часто
може здійснюватися кількома альтернативними шляхами. |
|
|
|||
Останнім етапом |
процесингу |
поліаденілуванняє |
3´-кінця |
іРНК – |
|
приєднання |
до 3´-кінцевого |
нуклеотиду |
іРНКpolyA-послідовності (100 |
– 200 |
|
аденінових |
нуклеотидів). Передбачається, що поліаденіловий «хвіст» забезпечує |
||||
транспорт |
зрілої іРНК |
до рибосоми, захищаючи її |
від ферментативного |
||
руйнування, але сам поступово руйнується ферментами цитоплазми, що відщеплюють один за одним кінцеві нуклеотиди іРНК.
Загальна схема будови зрілої еукаріотичної іРНК зображена на рис. 7.7. Між кепом і початком кодуючої ділянки(стартовим кодоном) розташована 5´-кінцева зона, яка не транслюється (5´UTR – untranslated region). За кодуючою ділянкою, яка закінчується одним із стоп-кодонів, і перед polyA-послідовністю міститься 3´- кінцева зона, що не піддається трансляції. Обидві ці зони містять важливі нуклеотидні послідовності, які використовуються для регуляції білкового синтезу.
Рис. 7.7. Схема будови зрілої еукаріотичної іРНК
Запитання для самоперевірки:
1.Що таке транскрипція? У чому полягає суть процесу транскрипції?
2.Якими ферментами забезпечується процес транскрипції?
3.Які ви знаєте основні стадії транскрипції?
4.Який ланцюг ДНК під час транскрипції називається кодуючим?
5.Охарактеризуйте будову транспортної РНК.
6.Як називають різні тРНК, які відповідають одній і тій же амінокислоті?
7.Від чого залежить порядок залучення амінокислот до поліпептидного ланцюга?
8.Від точності якого процесу буде залежати точність синтезу білка в цілому?
9.Охарактеризуйте будову рибосоми. Яка роль рибосомних білків?
10.Які ви знаєте активні центри рибосоми? Яке їх призначення?
11.Що таке трансляція? Назвіть та охарактеризуйте основні етапи трансляції.
12. Які |
ви |
знаєте |
особливості |
прокаріотичної |
системи |
транскр |
(трансляції)? |
|
|
|
|
|
|
13.Яким чином впізнається стартовий кодон прокаріотичною рибосомою?
14.Що значить полі- і моноцистронна іРНК?
15.Чи є принципові відмінності в організації прота еукаріотичного гену?
16.Що таке процесинг іРНК?
17.Яке функціональне значення кепу еукаріотичної іРНК?
18.Яка роль сплайсосоми у процесингу іРНК?
19.Яке призначення поліаденілового «хвоста» іРНК?
20.Охарактеризуйте загальну схему будови зрілої еукаріотичної іРНК. Які вона має відмінності у порівнянні з прокаріотичною іРНК?
64
ТЕМА 8. ЗАКОНИ МЕНДЕЛЯ
8.1. Гібридологічний аналіз
Одним із головних понять генетики є поняття ознаки– певної визначеної властивості організму або його частини, за якою одна особина відрізняється від іншої. Ознакою в генетичному розумінні можна вважати будь-яку характеристику на рівні зовнішніх рис, фізіологічних або біохімічних особливостей: вага, колір, ріст, набір речовин певного типу в органах і клітинах, сукупність певних білків тощо. Сукупність ознак даного організму називають фенотипом. Мендель усвідомив, що ознаки, які можна використати в генетичному аналізі, повинні виявлятися постійно (незалежно від віку, зовнішніх умов тощо) і мати два або більше альтернативних проявів.
Найважливішим експериментальним підходом генетики (практично єдиним
на початковому |
етапі |
її розвитку) є схрещування – природне або |
штучне |
поєднання двох |
гамет |
при заплідненні. Схрещування позначають |
знаком |
множення "´". У схемах на перше місце ставлять генотип жіночої ,статіяку позначать символом "♀" (дзеркало Венери), на друге – чоловічої із символом "♂" (щит і спис Марса). Під генотипом у схемах схрещувань розуміють не сукупність усіх генів організму, а тільки тих, що стосуються ознак, які аналізуються.
Домінантні та рецесивні алелі одного гена позначають, як правило, однаковими великою та малою літерами відповідно. Потомство від схрещування двох особин із різними альтернативними проявами ознаки називають гібридним, а окремого представника такого потомства– гібридом. Батьківські організми, які беруть у схрещування, позначають літерою Р (від латинського parentes – батьки), а потомство – літерою F (від латинського filii – діти). Зазвичай біля літери F
ставлять цифровий індекс, який відповідає порядковому номеру гібридного покоління.
Грегор Мендель відкрив закономірності успадкування, проводячи гібридизацію різних сортів гороху. Гібридизація - це схрещування особин з різними генотипами. Схрещування, при якому в батьківських особин враховується одна пара альтернативних ознак, називається моногібридним, дві пари ознак-
дигібридним, більш двох пар - полігібридним.
Гібридологічний метод Г. Менделя має наступні особливості:
·аналіз починається зі схрещування гомозиготних особин («чисті лінії»);
·аналізуються окремі альтернативні (взаємовиключні) ознаки;
·проводиться точний кількісний облік нащадків з різною комбінацією ознак (використовуються математичні методи);
·спадкування аналізованих ознак простежується в ряді поколінь.
8.2.Моногібридні схрещування (перший та другий закони Менделя)
Перший закон Менделя - закон однаковості гібридів першого покоління.
Г. Мендель схрещував чисті лінії рослин гороху з жовтим і зеленим насінням
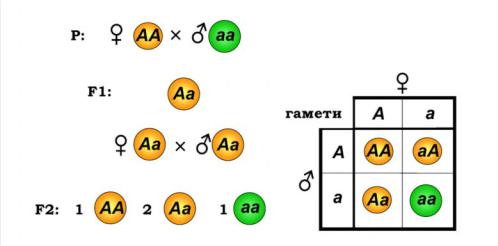
65
(альтернативні ознаки). Чисті лінії - це організми, що не дають розщеплення при схрещуванні з такими ж по генотипу, тобто вони є гомозиготними по даній ознаці. При аналізі результатів схрещування виявилося, що всі нащадки (гібриди)
у першому поколінні однакові по фенотипі(усі рослини мали горошини жовтого кольору) і по генотипі (гетерозиготи, рис. 8.1).
Рис. 8.1. Схема моногібридного схрещування (праворуч показано решітку Пеннета для другого схрещування)
Перший закон Менделя формулюється в такий спосіб: при схрещуванні |
|
|||||
гомозиготних особин, аналізованих по одній парі альтернативних ознак, |
|
|||||
спостерігається однаковість гібридів першого покоління як по фенотипу, так |
|
|||||
і по генотипу. |
|
|
|
|
|
|
Гібриди першого покоління продукують уже два типи гамет– з алелями А |
|
|||||
або а. При схрещуванні цих гібридів мають утворитися три різні комбінації алелів |
|
|||||
(строго кажучи, чотири комбінації, дві з яких – аА та Аа – є еквівалентними). Для |
|
|||||
полегшення |
розрахунків |
цих |
комбінацій |
іноді |
застосовують , |
таб |
запропоновану Пеннетом (її ще називають решіткою Пеннета): у першому рядку |
|
|||||
та першому стовпчику записують типи гамет, і тоді на перетині отримують |
|
|||||
комбінації алелів у нащадків. Із решітки Пеннета на рис. 8.1 видно, що частота |
|
|||||
гетерозигот Аа буде у два рази вищою, ніж однакові частоти гомозигот за |
|
|||||
домінантним і рецесивним алелями. Отже, для великої кількості особин другого |
|
|||||
покоління генотипи АА, Аа, |
аа будуть |
отримані у |
співвідношенні1 : 2 : 1, а |
|
||
співвідношення фенотипів (жовтих до зелених) буде дорівнювати 3 : 1. Саме цей |
|
|||||
результат і отримав Мендель. Тому другий закон Менделя - закон розщеплення. |
|
|||||
Особи, що містять домінантний ген А, мають жовте забарвлення насінь, а ті, |
|
|||||
що мають |
обидва рецесивних- зелене. |
Отже, відношення особин по фенотипу |
|
|||
(забарвленню насіння) - 3:1 (3 частини з домінантною ознакою і1 частина - з |
|
|||||
рецесивною). По генотипу: 1 частина особин – жовті гомозиготи (АА), 2 частини |
|
|||||
- жовті гетерозиготи(Аа) і 1 |
частина - зелені гомозиготи(аа). Другий закон |
Менделя формулюється так: |
при схрещуванні гібридів першого покоління |
(гетерозиготних організмів), |
аналізованих по одній парі альтернативних |
ознак, спостерігається розщеплення в співвідношенні 3:1 по фенотипу і 1:2:1 по генотипу.
66
Отримані групи нащадків Мендель піддав самозапиленню та встановив, що рослини з рецесивними ознаками не дають розщеплення в наступних поколіннях. Таким самим чином поводить себе й1/3 всіх особин із домінантною ознакою. А решта 2/3 домінантних особин дають розщеплення на домінантних і рецесивних знову ж таки у співвідношенні3 : 1. Отже, Мендель практично описав поведінку хромосом під час мейозу та запліднення. Важливість його роботи полягала ще й у демонстрації того, що, слідкуючи за спадкуванням зовнішніх ознак при схрещуваннях, можна робити висновки щодо генотипу особини.
8.3. Дигібридні і полігібридні схрещування (третій закон Менделя)
Вивчивши успадкування однієї пари алелей, Мендель вирішив простежити успадкування двох ознак одночасно. Для цієї мети він використав гомозиготні рослини гороху, що відрізняються по двох парах альтернативних ознак: насіння жовті гладкі і зелені зморшкуваті. У результаті такого схрещування у першому поколінні він одержав рослини з жовтим гладким насінням.
Цей результат показав, що закон однаковості гібридів першого покоління виявляється не тільки при моногібридному, але і при полігібридному схрещуванні, якщо батьківські форми гомозиготні. Потім він схрестив гібриди
першого |
покоління |
між |
собою. Для |
аналізу |
результатів |
полігібридного |
схрещування звичайно |
використовують |
решітку Пеннета(рис. 8.1): у |
першому |
|||
рядку та першому стовпчику записують типи гамет, і тоді на перетині отримують комбінації алелів у нащадків. У результаті вільного комбінування гамет у зиготи попадають гени в різних комбінаціях. Легко підрахувати, що по фенотипу потомство поділяється на4 групи: 9 частин рослин з горошинами жовтими гладкими (А-В-), 3 частини - з жовтими зморшкуватими(А-вв), 3 частини - із зеленими гладкими (ааВ-), 1 частина - із зеленими зморшкуватими(аавв), тобто розщеплення 9:3:3:1, чи (3+1)n. Звідси можна зробити висновок, що при схрещуванні гетерозиготних особин, що відрізняються по кількох парах альтернативних ознак, у потомстві спостерігається розщеплення по фенотипу в співвідношенні (3+1)n, де n- число ознак у гетерозиготному стані.
Якщо припустити (що й зробив Мендель), що два гени спадкуються незалежно, то дигібридні схрещування можна розглядати як два незалежні моногібридні схрещування. Якщо проаналізувати розщеплення по кожній з пар альтернативних ознак (жовтий і зелений колір, гладка і зморшкувата поверхня), то
вийде: 9+3 жовтих і 3 + 1 |
зелених, співвідношення 12:4 чи 3:1. Отже, при |
|
дигібридному |
схрещуванні |
кожна пара ознак у потомстві дає розщепленн |
незалежно від іншої пари. Це є результатом випадкового комбінування генів(і |
||
відповідних |
їм ознак), що |
приводить до нових сполучень, яких не було у |
батьківських формах. У нашому прикладі вихідні форми гороху мали насіння жовте гладке і зелене зморшкувате, а у другому поколінні отримали не тільки таке сполучення ознак, як у батьків, але і рослини з жовтим зморшкуватим і зеленим гладким насінням. Звідси випливає третій закон Менделя- закон незалежного комбінування ознак – при схрещуванні гомозиготних організмів, аналізованих
67
по двох (чи більше) парах альтернативних ознак, у другому поколінні спостерігається незалежне комбінування ознак і відповідних їм генів різних алельних пар. Це обумовлено генним рівнем організації спадкового матеріалу.
Для пояснення результатів схрещування, проведеного Г. Менделем, У. Бетсон (1902 р.) запропонував гіпотезу «чистоти гамет». Її можна звести до двох основних положень:
·у гібридному організмі гени не гібридизуються(не змішуються), а знаходяться в чистому алельному стані;
·внаслідок розбіжності гомологічних хромосом і хроматид при мейозі з кожної пари алелей у гамету потрапляє тільки один ген.
8.4.Умови для прояву законів Менделя
Закони Менделя носять статистичний характер(справедливі для великої кількості особин) і є універсальними, тобто при статевому розмноженні вони притаманні уcім живим організмам. Для прояву законів Менделянеобхідно дотримуватися ряду умов:
·гени різних алельних пар повинні знаходитися в різних хромосомах;
·між генами не повинне бути зчеплення і взаємод(крімї повного домінування);
·повинна бути рівна імовірність утворення гамет і зигот різного типу і рівна імовірність виживання організмів з різними генотипами(не повинно бути летальних генів);
·повинна бути 100% пенетрантність гена, відсутня плейотропна дія і мутації гена.
8.5.Причини відхилень від менделівських заномірностей спадкування ознак
З цитологічних основ спадковості, менделівські розщеплення у принципі можуть бути реалізованими тільки для видів із нормальним статевим процесом, диплоїдністю обох статей та за умови відсутності порушень у мейозі. Але й при виконанні зазначених умов випадки відхилень від менделівських розщеплень досить численні. При цьому менделівські закономірності не порушуються– просто на них накладаються інші ефекти.
Перш ніж з'ясовувати причини та механізми відхилень, у кожному конкретному випадку слід відповісти на питання, чи дійсно має місце таке відхилення від очікуваного за Менделем розподілу ознак. Адже відсутність відповідності між очікуваним та реальним співвідношенням фенотипів може бути викликана просто випадковими відхиленнями. Випадкові відхилення можуть приводити до різниці між теоретично очікуваними розщепленнями та такими, що спостерігаються в досліді. Чим більший розмір вибірки, тим меншими будуть такі відхилення. Якщо експериментальні дані відрізняються від теоретично очікуваних тільки за рахунок випадкових відхилень, то кажуть, що виправдовується нульова гіпотеза.
68
Відхилення від очікуваного розщеплення за законами Менде викликають:
·взаємодія неалельних генів;
· |
зчеплення груп генів в одній хромосомі та розташування гена |
|
статевих хромосомах; |
·летальність певних комбінацій алелів;
·існують алелі, які фенотипово виявляються тільки в певної частини організмів, що містять їх у генотипі. Для характеристики такого варіабельного прояву генів використовують поняття пенетрантності та експресивності. Мірою пенетрантності є частка , особинякі характеризуються певним фенотипом, серед усіх особин з однаковим генотипом. Деякі спадкові хвороби людини, наприклад подагра, характеризуються неповною пенетрантністю, тобто не всі особи, які несуть мутацію, хворіють. Експресивність відображає силу прояву мутантного гена у фенотипі. Так, дрозофіли, гомозиготні за мутантним алелем eyeless, можуть мати різну кількість фасеток очей– від нормальної кількості до повної їхньої відсутності. Здатність генотипу виявлятися по-різному залежно від зовнішніх умов відображає його норму реакції – певний діапазон прояву, у межах якого ознаки можуть варіювати у відповідь на варіації умов розвитку організму;
·деякі ознаки виникають лише в певний період: характер розщеплення може залежати від віку. Наприклад, у людини є спадкові синдроми, які виявляються лише в похилому віці(синдром Альцгеймера, хвороба Гентінгтона);
· |
до відхилень від очікуваних менделівських розщеплень приводить |
|
також явище імпринтингу. Основою його є так звана епігенетична |
|
спадковість – успадкування від батьків не просто ДНК, а хроматину, |
|
який несе на собі певні хімічні маркери, від яких залежить структурний |
|
стан ділянки та, відповідно, активність певних генів. Унаслідок такого |
|
ефекту прояв певного гена може бути зумовлений тим, від кого саме з |
|
батьків дана хромосома отримана; |
Запитання для самоперевірки:
1.Що таке гібридизація?
2.Які схрещування називаються моногібридними?
3.Назвіть особливості гібридологічного методу Г. Менделя.
4.Сформулюйте перший закон Г. Менделя.
5.Поясніть другий закон Г. Менделя.
6.Чому третій закон Г. Менделя має назву закону незалежного комбінування ознак?
7.Назвіть положення гіпотези чистоти гамет.
8.Яких умов необхідно дотримуватись для прояву законів Г. Менделя?
9.Назвіть можливі причини відхилень від менделівських заномірностей спадкування ознак.
69
ТЕМА 9. ВЗАЄМОДІЯ ГЕНІВ
9.1. Взаємодія алельних генів: повне домінування, неповне домінування, наддомінування, кодомінування
Взаємодія алельних генів називаєтьсявнутрішньоалельною. Виділяють наступні його види:
·повне домінування;
·неповне домінування;
·наддомінування;
·кодомінування.
При повному домінуванні один ген цілком пригнічує прояв іншого гена (виконуються закони Менделя), при цьому гомозиготи по домінантній ознаці і гетерозиготи фенотипово нерозрізняються. Наприклад, ген жовтого кольору насіння гороху цілком пригнічує ген зеленого забарвлення, ген карого кольору очей у людини пригнічує ген блакитного забарвлення.
При неповному домінуванні (проміжному спадкуванні) домінантний ген не повністю пригнічує прояв дії рецесивного гена. У гібридів першого покоління спостерігається проміжне успадкування, а в другому поколінні - розщеплення по фенотипу і генотипу однаково- 1:2:1 (виявляється доза дії генів). Наприклад, якщо схрестити рослини запашного горошку з червоними і білими квітами перше покоління буде мати рожеві квітки.
При наддомінуванні домінантний ген у гетерозиготному стані виявляє себе сильніше, ніж у гомозиготному. У мухи дрозофіли є рецесивний летальний ген (а) - гомозиготи (аа) гинуть. Мухи, гомозиготні по гену А(АА) мають нормальну
життєздатність, а гетерозиготи (Аа) |
- |
живуть довше і більш плідні, чим |
|
домінантні гомозиготи. Пояснити |
це |
можна взаємодією |
продуктів генної |
активності. |
|
|
|
При кодомінуванні гени однієї алельної пари рівнозначні, |
жоден з них не |
||
пригнічує дії іншого; якщо вони обидва знаходяться в генотипі, обидва виявляють свою дію. Типовим прикладом кодомінування є успадкування груп крові людини
по АВО- |
(група |
АВ) |
і МN- (група МN) |
системах. Одночасна присутність у |
||
генотипі |
генів JA |
і JB |
обумовлює |
наявність в еритроцитах антигенів А і (IVВ |
||
група крові). Гени JA і JB не пригнічують один одного- вони є рівноцінними, |
||||||
кодомінантними. |
|
|
|
|
|
|
9.2. Взаємодія неалельних генів: компліментарність, епістаз, полімерія |
||||||
Взаємодія |
неалельних |
генів |
називаєтьсяміжалельною. |
Розрізняють |
||
наступні його види:
·комплементарність;
·епістаз;
·полімерія.
При комплементарності присутність в одному генотипі двох домінантних
70
(рецесивних) генів з різних алельних пар приводить до появи нового варіанта ознаки. Типовий приклад - розвиток слуху людини. Для нормального слуху в генотипі людини повинні бути присутні домінантні гени з різних алельних пар–
D і Е. Ген D відповідає за нормальний розвиток равлика вуха, ген Е– за розвиток слухового нерва. У рецесивних гомозигот(dd) буде недорозвинений равлик вуха, а при генопі ее - недорозвинений слуховой нерв. Люди з генотипами D-ее, ddЕ- і ddее будуть глухими.
При епістазі домінантний (рецесивний) ген з однієї алельної пари пригнічує дію домінантного (рецесивного) гена з іншої алельної пари. Це явище протилежне комплементарності. Пригнічуючий ген називається супресором(інгібітором). У курей домінантний ген С детермінує синтез пігменту, а домінантний алель іншого гена I є його супресором, кури з генотипом С-І- мають біле пір’я.
При полімерії домінантні гени з різних алельних пар впливають на ступінь прояву тієї самої ознаки. Полімерні гени прийнято позначати однією буквою латинського алфавіту з цифровими індексами, наприклад, А1А1, А2А2 а3а3, і т.д. Ознаки, які детермінуються полімерними генами, називаються полігенними. У такий спосіб успадковуються багато кількісних і деяких якісних ознак у тварин і людини: ріст, маса тіла, величина артеріального тиску, колір шкіри й ін. Ступінь прояву цих ознак залежить від кількості домінантних генів у генотипі(чим їх більше, тим сильніше виражена ознака) і значною мірою від впливу умов середовища. У людини може спостерігатися схильність до різних захворювань: гіпертонічної хвороби, ожирінню, цукровому діабету, шизофренії й ін. Дані
ознаки |
при сприятливих умовах середовища можуть і не проявитися чи бути |
слабко |
вираженими. Це відрізняє полігенно наслідувані ознаки від моногенних. |
Змінюючи умови середовища і проводячи профілактичні заходи можна значно |
|
знизити частоту і ступінь виразності деяких мультифакторних захворювань. |
|
Підсумовування «доз» |
полімерних генів (адитивна дія) і вплив середовища |
|||
забезпечує існування неперервних рядів кількісних змін. Мінімальна кількість |
|
|||
полімерних |
генів, при |
якому виявляється |
ознака, називається граничним |
|
ефектом. |
|
|
|
|
|
|
Запитання для самоперевірки: |
|
|
1. Що |
таке |
внутрішьоалельна |
взаємодія? Якігенів є |
види |
внутрішньоалельної взаємодії генів? |
|
|
||
2.Наведіть приклад повного домінування генів.
3.Охарактеризуйте явище неповного домінування генів.
4.Поясніть явище наддомінування генів.
5.За якого типу взаємодії алельних генів різні гени однієї алельної пари проявляються у фенотипі?
6.Які ви знаєте види взаємодій неалельних генів?
7.Поясніть комплементарну взаємодію генів.
8.Як проявляється епістатична взаємодія генів?
9.Охарактеризуйте явище полімерії. Як називають полімерні гени?