

9.11. Ейкозаноїди
Ейкозаноїди – це біологічно активні речовини, які синтезуються в багатьох клітинах із полієнових жирних кислот, що містять 20 атомів карбону («ейкоза» – 20, грец). Вони включають у себе простагландини, тромбоксани та лейкотрієни тощо – високоактивні регулятори клітинних функцій. Їх період півжиття надзвичайно короткий, тому свої ефекти вони чинять як «гормони місцевої дії», впливаючи на метаболізм тих клітин, що їх синтезують за автокринним механізмом, а на оточуючі клітини – за паракринним механізмом.
В
Рис.
9.15. Вивільнення арахідонової кислоти
із мембранних ліпідів: зв’язування
активоторів із рецепторами плазматичної
мембрани активує шлях 1 або 2: ДАГ-ліпаза
– діацилгліцеролліпаза, МАГ –
моноацилгліцеролліпаза
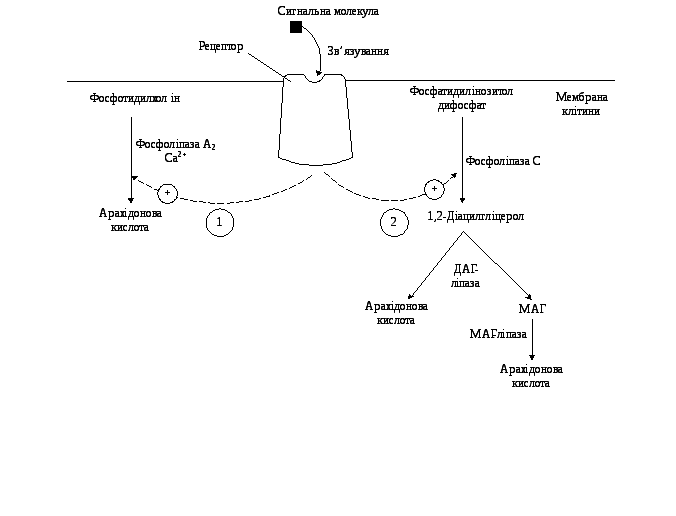
Головними субстратами для синтезу ейкозаноїдів слугують арахідонова, ейкозапентаєнова та ейкозатрієнова кислоти. Вони входять до складу гліцерофосфоліпідів мембран (рис. 9.15).
Під впливом асоційованої з мембраною фосфоліпази А2 (рідше фосфоліпази С) жирна кислота відщеплюється від гліцерофосфоліпіда та використовується для синтезу ейкозаноїдів. У різних тканинах арахідонова та інші ейкозанові кислоти можуть використовуватися в трьох основних напрямках: циклооксигеназний шлях призводить до утворення простагландинів, простациклінів і тромбоксанів; ліпооксигеназний шлях забезпечує синтез лейкотрієнів, ліпоксинів і гідроксіейкозатетраєноатів (ГЕТЄ); система окиснення за участі цитохрому Р-450 бере участь у синтезі епоксидів.
9
Рис.
9.16. Циклооксигеназний шлях перетворення
арахідонової кислоти. Знаком „-”
позначено гальмівну дію вказаних
речовин
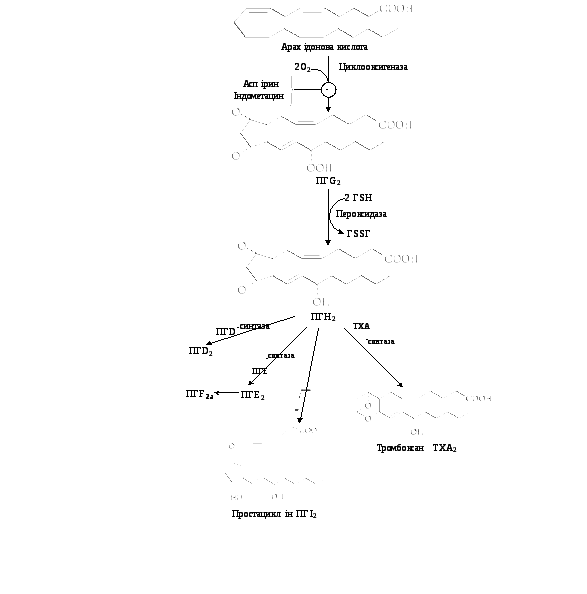
Біосинтез ПГ і тромбоксану починається із перетворення арахідонової кислоти на ендопероксидні проміжні продукти (ПГG2 і ПГН2) (рис. 9.16). Обидві реакції каталізуються поліферментним комплексом простагландинсинтазою, який складається із двох компонентів — циклооксигенази (ЦОГ) і пероксидази. В активному центрі ЦОГ міститься тирозин, а пероксидази – гем. В організмі присутні два типи ЦОГ: ЦОГ-1 конститутивний фермент, який синтезується з постійною швидкістю, тоді як синтез ЦОГ-2 зростає під час запалення та індукується відповідними медіаторами – цитокінами.
Обидва типи циклооксигеназ каталізують включення 4 атомів кисню в арахідонову кислоту з утворенням п’ятичленного кільця. До утворення ПГН2 шлях синтезу різних ПГ однаковий, подальші перетворення специфічні для кожного типу клітин. так, у тромбоцитах під впливом тромбоксансинтази з ПГН2 утворюється тромбоксан А2 (ТхА2), який володіє потужною судинозвужувальною дією, а в клітинах ендотелію – простациклін (ПГІ2), який розширює судини.
9.11.2. Біосинтез лейкотрієнів. Лейкотрієни також синтезуються з ейкозанових кислот, однак, на відміну від ПГ, у їх структурі відсутні цикли і вони мають 3 спряжені подвійні зв’язки (звідки назва «лейкотрієн»).
Утворення лейкотрієнів відбувається переважно у клітинах крові – лейкоцитах різних класів, тромбоцитах, макрофагах, що відображає провідну роль лейкотрієнів у реакціях запалення, згортанні крові, алергійних реакціях і імунних процесах.
П
Рис.
9.17. Схема синтезу лейкотрієнів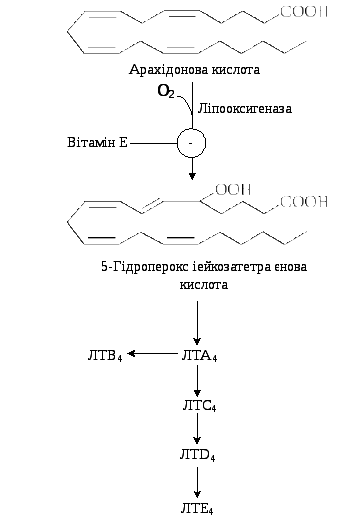
Синтезовані ейкозаноїди надходять із клітин у міжклітинне середовище і реалізують свої функції, взаємодіючи із специфічними рецепторами мембран клітин-мішеней. Інактивація цих речовин відбувається дуже швидко (від кількох секунд до кількох хвилин) під дією відповідних ферментів. Продукти перетворень ейкозаноїдів виводяться з сечею.
9.11.3. Основні біологічні ефекти ейкозаноїдів. Всі ейкозаноїди, незважаючи на короткий період піврозпаду, мають надзвичайно широкий спектр дії та високу біологічну активність. Вони діють на клітини через спеціальні рецептори. Деякі з них зв’язані з аденілатциклазною системою та протеїнкіназою А (ПГЕ, ПГD, ПГІ2); інші діють через зростання рівня кальцію в цитозолі клітин мішеней (ПГF, ТхА2, лейкотрієни). У різних тканинах вони виконують різні, іноді протилежні функції (табл. 9.4).
Таблиця 9.4. Біологічні функції ейкозаноїдів
|
Ейкозаноїди |
Локалізація |
Біологічна активність |
|
ПГЕ2 |
Більшість тканин, особливо нирки |
Розширення судин, розслаблення гладких м’язів, стимуляція пологової діяльності, пригнічення міграції лімфоцитів, проліферації Т-клітин, агрегація тромбоцитів |
|
ПГF2 |
Більшість тканин |
Звуження судин, бронхо – і вазоконстрикція, скорочення гладких м'язів |
|
ПГD3 |
Клітини гладких м’язів |
Розширення судин, зниження агрегації тромбоцитів і лейкоцитів |
|
ПГІ2 |
Серце, ендотелій судин |
Розширює судини, попереджує агрегацію тромбоцитів, підвищує рівень цАМФ у клітині |
|
ТХА2 |
Тромбоцити |
Стимулює агрегацію тромбоцитів, звужує судини і бронхи, у клітинах знижує утворення цАМФ |
|
ЛТВ4 |
Моноцити, базофіли, нейтрофіли еозинофіли, епітеліальні клітини |
Індукує хемотаксис і агрегацію лейкоцитів, вивільнення лізосомальних ферментів лейкоцитів, посилює проникність судин |
|
ЛТС4, ЛТD4, ЛТЕ4 |
Лейкоцити, макрофаги |
Розширюють судини і збільшують їх проникність, викликають спазм бронхів, є компонентами "повільно реагуючої" субстанції анафілаксії |
Біологічні функції ПГ пов'язані з впливом на скорочувальну функцію гладких м'язів, проте окремі ПГ мають різні фізіологічні ефекти у певних тканинах-мішенях. Так, наприклад, ПГ А та Е знижують артеріальний тиск при гіпертонічній хворобі; ПГЕ спричинює розслаблення гладеньких м'язів бронхів і трахеї, тоді як ПГF, навпаки, викликає їх скорочення.
Характерною особливістю ПГЕ2 та ПГF2 (містяться в сім'яній рідині) є їх стимулюючий вплив стосовно м'язів матки, що сприяє переміщенню сперматозоїдів у порожнину фаллопієвих труб, де відбувається запліднення. ПГ Е1 гальмує базальну та стимульовану секрецію хлоридної кислоти, захищає клітини слизової оболонки від ушкоджуючих хімічних подразників. Препарати ПГЕ1 мізопростол, цитотек є ефективними засобами сучасної терапії виразкової хвороби.
Протилежну дію мають простациклін і тромбоксан. Простациклін синтезується у клітинах ендотелію судин і перешкоджає згортанню крові, гальмуючи агрегацію тромбоцитів і розширюючи судини, а тромбоксан А2 утворюється у тромбоцитах і сприяє їх агрегації, а також скорочує гладкі м'язи судин. Звільнення тромбоксану із агрегатів тромбоцитів і дифузія в стінку судин зумовлюють їх звуження. Таким чином, відносна активність тромбоксану і простацикліну визначає ймовірність утворення тромбів і місцевого спазму судин. Дисбаланс цих сполук у бік тромбоксанів відіграє важливу роль в утворенні атеросклеротичних бляшок. Механізм дії тромбоксану полягає у підвищенні виходу Са2+ із внутрішньоклітинних депо в цитозоль тромбоцитів. Іони кальцію, в свою чергу, стимулюють скорочувальні білки тромбоцитів, а також вивільнення із тромбоцитів вмісту їх гранул (серотоніну, катехоламінів, АДФ). Простациклін підвищує рівень цАМФ у тромбоцитах, що перешкоджає мобілізації Са2+ і синтезу тромбоксану. Простагландини також можуть впливати на рівень цАМФ у клітинах, зокрема простагландини групи Е активують аденілатциклазу в ендокринних залозах, але гальмують у жировій тканині. Концентрація цАМФ у клітині, своєю чергою, впливає на синтез ПГ.
Л
Рис.
9.18.
Механізм інактивації циклооксигенази
аспірином: ацетильний залишок переноситься
з молекули аспірину на ОН-групу фермента
та незворотно інгібує її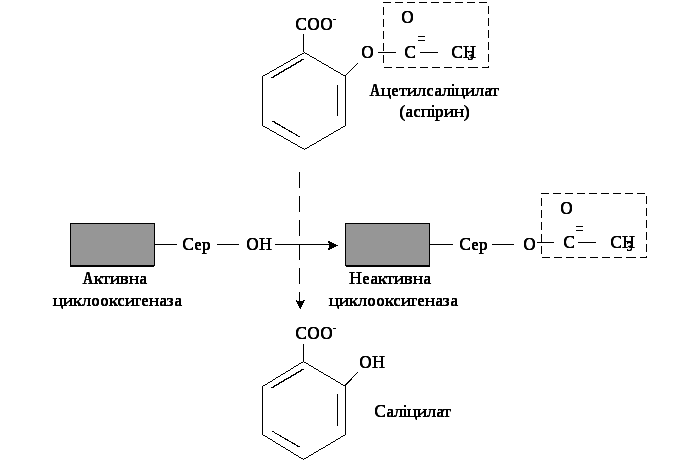
Аспірин – протизапальний препарат, механізм дії якого полягає в інгібуванні циклооксигенази і, як наслідок, пригніченні синтезу медіаторів запалення. Циклооксигеназа незворотно інгібується шляхом ацетилування серину в активному центрі фермента (рис. 9.18). Однак, ефект аспірину нетривалий, оскільки експресія гена цього фермента не порушується і синтезуються його нові молекули. Інші нестероїдні препарати (ібупрофен, ацетамінофен тощо) теж знижують синтез ПГ за конкурентним механізмом, зв’язуючись з активним центром фермента. Стероїдні препарати володіють значно потужнішою протизапальною дією, оскільки індукують синтез білків-ліпокортинів, котрі інгібують активність фосфоліпази А2 і, запобігаючи вивільненню субстрату для синтезу ейкозаноїдів – арахідонової кислоти, зменшують їх синтез.