Билет 30
Органы вкуса и обоняния. Строение, топография, кровоснабжение, иннервация.
У человека орган обоняния, orgdnum olfactorium, располагается в верхнем отделе носовой полости. Обонятельная область слизистой оболочки носа, regio olfactoria tunicae mucosae nasi, включает слизистую оболочку, покрывающую верхнюю носовую раковину и верхнюю часть перегородки носа. Рецепторный слой слизистой оболочки представлен обонятельными нейросенсорными клетками cellulae neurosensoriae olfactoriae, воспринимающими присутствие пахучих веществ. Под обонятельными клетками лежат поддерживающие клетки, cellulae sustentaculares. В слизистой оболочке находятся обонятельные железы, glandulae olfactoriae, секрет которых увлажняет поверхность рецепторного слоя. Периферические отростки обонятельных клеток несут на себе обонятельные волоски (реснички), а центральные формируют обонятельные нервы, nn. olfactorii. Обонятельные нервы через отверстия решетчатой пластинки одноименной кости проникают в полость черепа, затем в обонятельную луковицу, где аксоны обонятельных нейросенсорных клеток в обонятельных клубочках вступают в контакт с митральными клетками. Отростки митральных клеток в толще обонятельного тракта направляются в обонятельный треугольник, а затем в составе обонятельных полосок (промежуточной и медиальной) вступают в переднее продырявленное вещество, в подмозолистое поле, area subcallosa, и диагональную полоску, bandaletta diagonalis. В составе латеральной полоски отростки митральных клеток следуют в парагиппокампальную извилину и в крючок, в котором находится корковый центр обоняния.
Орган вкуса, orgdnum giistus.
У человека вкусовые почки, саlliculi gustatorii находятся в слизистой оболочке языка, а также неба, зева, надгортанника. Наибольшее количество вкусовых почек сосредоточено в желобоватых, papillae vallatae, и листовидных сосочках, papillae foliatae, меньше их в грибовидных сосочках, papillae fungiformes, слизистой оболочки спинки языка. В нитевидных сосочках их не бывает вообще. Каждая вкусовая почка состоит из вкусовых и поддерживающих клеток. На вершине почки имеется вкусовое отверстие (пора), porus gustatorius, открывающееся на поверхность слизистой оболочки.
На поверхности вкусовых клеток располагаются окончания нервных волокон, воспринимающих вкусовую чувствительность. В области передних 2/з языка это чувство вкуса воспринимается волокнами барабанной струны лицевого нерва, в задней трети языка и в области желобоватых сосочков — окончаниями языко-глоточного нерва. Этот нерв осуществляет вкусовую иннервацию также слизистой оболочки мягкого неба и небных дужек. От редко расположенных вкусовых луковиц в слизистой оболочке надгортанника и внутренней поверхности черпаловидных хрящей вкусовые импульсы поступают через верхний гортанный нерв — ветвь блуждающего нерва. Центральные отростки нейронов, осуществляющих вкусовую иннервацию в полости рта, направляются в составе соответствующих черепных нервов (VII, IX, X) к общему для них чувствительному ядру, nucleus solitarius, лежащему в задней части продолговатого мозга. Аксоны клеток этого ядра направляются в таламус, где импульс передается на следующие нейроны, заканчивающиеся в коре большого мозга, крючка парагиппокампальной извилины. В этой извилине находится конец вкусового анализатора.
Билет 31
Парасимпатический отдел вегетативной нервной системы. Общая характеристика; центры и периферическая часть (узлы, распределение ветвей).
Парасимпатическая часть, pars parasympathica (parasympathetica),автономной (вегетативной) нервной системы подразделяется на головной и крестцовый отделы. К головному отделу [pars cranidlis] относятся вегетативные ядра и парасимпатические волокна глазодвигательного (III пара), лицевого (точнее, промежуточного, — VIII пара), языкоглоточного (IX паpa) и блуждающего (X пара) нервов, а также ресничный, крылонебный, поднижнечелюстной, подъязычный и ушной узлы и их ветви. Крестцовый отдел [pars pelvica] парасимпатической части представлен крестцовыми парасимпатическими ядрами, nuclei parasympathetici sacrales, II, III и IV крестцовых сегментов спинного мозга, внутренностными тазовыми нервами, пп. splanchnici pelvini, и парасимпатическими тазовыми узлами, ganglia pelvina, с их ветвями.
1. Парасимпатическая часть глазодвигательного нерва представлена добавочным (парасимпатическим) ядром, nucl. oculo-motorius accessorius, так называемым ядром Якубовича, ресничным узлом и отростками клеток, расположенных в этом ядре и узле. Аксоны клеток добавочного ядра глазодвигательного нерва, залегающего в покрышке среднего мозга, проходят в составе III пары черепных нервов в виде преганглионарных волокон.
2. Парасимпатическая часть лицевого нерва состоит из верхнего и слюноотделительного ядра, крылонебного, поднижнече-люстного и подъязычного вегетативных узлов. Аксоны клеток верхнего слюноотделительного ядра, лежащего в покрышке моста, проходят в составе лицевого (промежуточного) нерва в одноименном канале.
3. Парасимпатическая часть языкоглоточного нерва образована нижним слюноотделительным ядром, ушным узлом и отростками залегающих в них клеток. Аксоны клеток нижнего слюноотделительного ядра, находящегося в продолговатом мозге, в составе языкоглоточного нерва выходят из полости черепа через яремное отверстие.
4. Парасимпатическая часть блуждающего нерва состоит из заднего (парасимпатического) ядра блуждающего нерва, многочисленных узлов, входящих в состав органных вегетативных сплетений и отростков клеток, расположенных в ядре и этих узлах. Аксоны клеток заднего ядра блуждающего нерва, находящегося в продолговатом мозге, идут в составе ветвей блуждающего нерва. Они достигают парасимпатических узлов,ganglia parasympathica, околоорганных и внутриорганных вегетативных сплетений.
5.
Крестцовый отдел парасимпатической
части автономной (вегетативной)
нервной системы представлен крестцовыми
парасимпатическими ядрами, nuclei
parasympathetia sac-rales, расположенными в
латеральном промежуточном веществе 11
крестцовых сегментов спинного мозга,
тазовыми (парасимпатическими)
узлами,ganglia pelvina, и отростками залегающих
в них клеток. Аксоны клеток крестцовых
парасимпатических ядер выходят из
спинного мозга в составе передних
корешков, затем идут в составе передних
ветвей крестцовых спинномозговых нервов
и после выхода их через тазовые крестцовые
отверстия ответвляются, образуют тазовые
внутренностные нервы, пп. spldnchnici
pelvini.


Билет 32
Пирамидные пути.
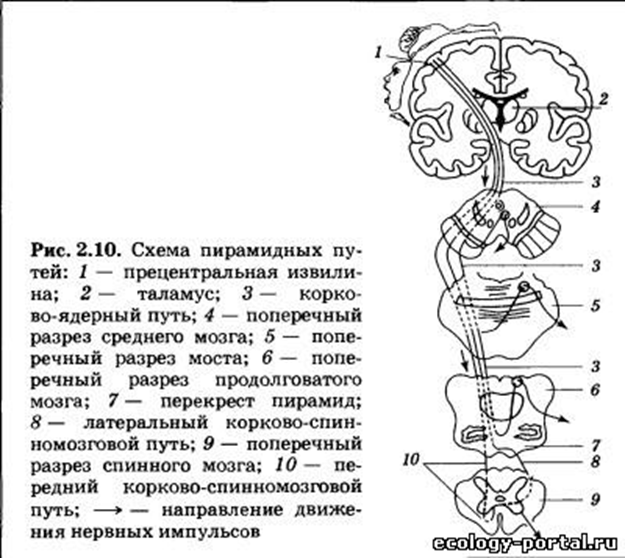
К пирамидному пути (tractus piramidalis) относится система волокон, по которым двигательные импульсы из коры большого могзга, из предцентральной извилины, гигантопирамидальных нейронов направляются к двигательным ядрам черепных нервов и передним рогам СМ, а от них к скелетным мышцам. Пирамидный путь подразделяется на 3 части: 1) корково-ядерный путь - к ядрам черепных нервов, 2) латеральный корково спинномозговой - к ядрам передних рогов СМ, 3) передний корково-спинномозговой - также к передним рогам СМ.
1. Корково-ядерный путь - пучок готростком гигантопирамидальных нейронов, которые из коры нижней трети предцентральной извилины спускаются к внутренне капсуле и проходят через ее колено. Затем идут в основании ножки мозга, образуя медиальную часть пирамидных путей. Начиная со среднего мозга и далее, в мосту и продолговатом мозге волокна корково-ядерного пути переходят на противоположную сторону к двигательным ядрам черепных нервов: 3 и 4 - в среднем мозге; 5,6,7 - в мосту; 9,10,11,12 - в продолговатом мозге. В этих ядрах корково-ядерный путь заканчивается. Отростки упомянутых двигательных клеток выходят из мозга и в составе черепных нервов идут к скелетным мышцам.
2. Латеральный и передний корково-спинномозговые пути также начинаются от гигантопирамидальных нейронов предцентральной извилины. Аксоны этих клеток идут к внутренней капсуле, проходят через переднюю часть ее задней ножки, спускаются в основание ножки мозга, где занимают место латеральнее корково-ядерного пути. Далее волокна спускаются в основание мозга, пронизывают пучки волокон моста и выходят в продолговатый мозг, где образуют пирамиды. В нижней част часть волокон переходит на противоположную сторону и продолжается в боковой канатик СМ, заканчиваясь в передних рогах СМ синапсами на двигательных клетках его ядер. Часть, которая участвует в образовании перекреста пирамид - латеральный корково-спинномозговой путь. Те волокна, которые в нем не участвуют и не перекрещиваются составляют передний корково-спинномозговой путь (затем они также переходят на противоположную сторону, но через белую спайку СМ и заканчиваются на двигательных клетках переднего рога противоположной стороны СМ).
Вторыми нейронами нисходящего корково-спинномозгового пути являются клетки передних рогов СМ, длинные отростки которых выходят из СМ в составе передних корешков и идут в составе спинномозговых нервов для иннервации скелетных мышц.
Билет 33
Понятие о нейроне (нейроците). Нервные волокна, корешки и пучки; межпозвоночные узлы, их классификация и строение.
Структурно-функциональной единицей и активным элементом нервной системы является нейрон с окружающей его глиальной тканью и кровеносными микрососудами. Глиальная ткань (глия) представляет собою специализированный вид соединительной ткани, обеспечивающий нейронам опорную, защитную и трофическую функции. Новейшие исследования на молекулярном уровне показывают присутствие в нервных клетках макромолекул белков, реагирующих на раздражитель. Подобные макромолекулы обнаружены и в других клетках – например в мышечных. Это доказывает: нейрон — не уникальная по строению клетка, а всего лишь специализировавшаяся на выполнении особых функций.
Нервная клетка обладает телом разнообразной формы, из которого вырастают два вида отростков: короткие (до нескольких мм) и чаще многочисленные дендриты; длинные (до 1 м) — чаще одиночные, нитевидные аксоны (нейриты) с многочисленными ответвлениями (коллатералями). Проведение возбуждения в нейроне строго поляризовано: от менее активного дендрита к более активному аксону. Однако, по современным представлениям нейроны могут получать дифференцированные сигналы, активно проводимые дендритной системой и выборочно передаваемые на аксоны в зависимости от источника, уровня и синхронности сигналов. На всех своих терминалях нейрон синтезирует и выделяет единственный медиатор или трансмиттер (закон Дейла) — так считалось ранее. Теперь же в каждом синапсе вместе с классическими медиаторами соседствуют в разных сочетаниях новые — различные нейропептиды: энкефалин, соматостатин, галанин, мотилин и многие другие, обеспечивая модуляцию синаптической передачи. Кроме того, нейроны синтезируют цитокины и нейротрофины, которые переносятся аксонами и влияют как на электрические свойства и эффективность синаптического переключения, так и на образование медиатора (трансмиттера).
Нейроны достаточно устойчивы к повреждению, особенно при гипотермии и блокаде возбудительных рецепторов во время вредоносного действия. В поврежденном нейроне в большинстве случаев происходит перестройка цитоскелета с восстановлением ветвления отростков и продукции медиатора. Структурное пластическое перестроение нервной клетки под действием афферентных сигналов и обучения в обогащенной среде обитания протекает гораздо быстрее, чем считалось ранее. Разрушению и гибели нервных клеток противостоят образование, пролиферация и дифференцировка новых нейронов из стволовых клеток. Процессы восстановления можно усиливать введением нейротрофических средств, пересадкой нервной ткани, а в перспективе клонированием клеток-предшественников с применением методов генной инженерии.
Нервные клетки подразделяются:
• на мультиполярные с большими телами многоугольной, неправильной формы, многочисленными, короткими дендритами и одним длинным аксоном (они располагаются в головном и спинном мозге);
• на биполярные с небольшими телами овальной формы и отростками: дендритом и аксоном, отходящими от противоположных концов тела; такие нейроны часто встречаются в периферических отделах нервной системы;
• на униполярные с телом округлой формы и одним длинным отростком, распадающимся вблизи тела на две ветви;
• на псевдоуниполярные нейроны, которые обладают двумя сросшимися отростками, от чего клетка превращается в ложноуниполярную — такие нейроциты занимают спинномозговые узлы и другие чувствительные узлы;
• на нейросекреторные клетки, выделяющие в кровь и по отросткам гормоны (нейросекрет), например, в гипоталамусе, гипофизе и эпифизе;
• на нейроглиальные клетки: эпендимоциты, астроциты, олигодендроциты, глиальные макрофаги, выполняющие в нервной системе опорную, разграничительную, иммунную, трофическую и другие функции;
• на стволовые клетки мозга, способные к самовосстановлению и формированию предшественников всех перечисленных выше клеток от нейронов до глиоцитов; на протяжении всей жизни предшественники мигрируют в различные области мозга, проходят дифференцировку, интегрируются в работающие системы мозга.
Кровеносные микрососуды — артериолы, прекапилляры, капилляры, посткапилляры и венулы — осуществляют питание нервных клеток и глиальной ткани. Однако, часть нейронов и глиальных клеток головного и спинного мозга питается и за счет ликвора, который циркулирует в желудочках и центральном канале, расположенных внутри мозга.
Между собой нервные клетки контактируют при помощи синапсов, которые в виде бляшек, диаметром до 1 мкм покрывают тело нейрона (аксосоматические контакты) и его дендриты (аксодендритические контакты). Внутри бляшки находится синаптическая щель и синаптический пузырек, заполненный медиатором (ацетилхолином, адреналином), синаптома. При прохождении импульса медиаторы выходят из пузырька в щель, замыкают контакт и разрушаются. В настоящее время выявлены и другие способы общения нейронов: воздействие молекул трансмиттеров на внесинаптические рецепторы, на щелевые контакты, на микроциркуляторное русло.
По структурно-функциональным возможностям нейроны подразделяются на следующие группы.
Чувствительные нервные клетки воспринимают раздражение, иначе называются афферентными нейронами. Тела таких псевдоуниполярных нейроцитов располагаются вне мозга — в периферических узлах (ганглиях) черепных и спинальных нервов. Длинную ветвь отростка они посылают в органы, где она заканчивается нервным окончанием (рецептором). Короткая ветвь уходит в мозг, где вступает в синаптический контакт с центральными нейронами.
Эффекторные, эфферентные (двигательные или секреторные нейроны) находятся в мозге или в вегетативных узлах. Аксоны уходят в органы и железы, где заканчиваются двигательными или секреторными окончаниями.
Замыкательные, вставочные или кондукторные, ассоциативные нейроны (более правильный и современный термин) присутствуют в большом количестве. Они передают импульсы от афферентных нейронов через синапсы к эфферентным (исполнительным) и многим другим нервным клеткам. Н. М. Бехтерева открыла ассоциативные нейроны, передающие информацию по избирательному смыслу.
Отростки нейронов (нервные волокна) в периферической системе образуют корешки, пучки, нервы и нервные сплетения. Главной частью нервного волокна является осевой цилиндр, представляющий короткий или длинный вырост цитоплазмы, окруженный внутренней оболочкой — неврилеммой. В зависимости от строения наружной оболочки, покрывающей осевые цилиндры с неврилеммой, различают два типа волокон.
• Мякотные или миелиновые волокна, которые содержат в наружной, шванновской оболочке миелин — химическое вещество липоидного характера. Такая оболочка чехлом окружает осевой цилиндр, но на равных промежутках прерывается перехватами Ранвье, а ее миелиновые сегменты пронизаны косыми насечками.
• Безмякотные, безмиелиновые волокна не содержат миелина в наружной оболочке. Осевые цилиндры в них окружены тонким и равномерным слоем шванновского синцития. Но осевые цилиндры могут терять оболочки (мякотную, безмякотную) и тогда цитоплазму окружает только неврилемма. Такие осевые цилиндры часто встречаются в терминалях нервных окончаний.
В периферических нервах волокна складываются в пучки и удерживаются в них благодаря наличию периневрия — соединительно-тканной оболочки, которая окружает пучок. Между пучками нервных волокон располагаются в клетчатке кровеносные сосуды, питающие нерв. Пучки объединяются в нерв при помощи общей фиброзной оболочки — эпиневрия.
В головном и спинном мозге дендриты и аксоны образуют для связи нервных клеток несколько волоконных систем.
• Ассоциативные волокна распространяются в пределах одного полушария или половины спинного мозга.
• Комиссуральные волокна связывают между собой полушария или правую и левую половины спинного мозга.
• Проекционные волокна: восходящие и нисходящие проходят между выше и нижележащими отделами мозга и органами.
Корешки — нервные волокна, расположенные на основании головного мозга и в латеральных бороздах (передней и задней) спинного мозга. Они образуются:
• длинными отростками мозговых мотонейронов и называются двигательными, в спинном мозге они всегда передние;
• или образуются отростками псевдоуниполярных клеток спинальных и черепных узлов и называются чувствительными;
• а если они возникают из парасимпатических и симпатических узлов, то называются вегетативными.
В вегетативной системе волокна делят по отношению к периферическим ганглиям.
Преганглионарные волокна, которые покрыты миелиновой оболочкой и для передачи сигнала как в симпатической, так и в парасимпатической системе используют ацетилхолин. Поэтому их называют холинэргическими.
Постганглионарные волокна — безмиелиновые. В симпатической системе используют в качестве посредника в передаче импульса адреналин — адренэргические волокна. В парасимпатической системе — ацетилхолин (холинэргические волокна).
По морфо-функциональным параметрам волокна классифицируют в зависимости от величины диаметра и скорости проведения импульса. Чем толще волокно, тем быстрее проходит импульс.
Нервные корешки — черепные, спинномозговые, вегетативные, сливаясь вместе, образуют периферические нервы: 12 пар черепных и 31 пару спинномозговых. По составу волокон — чувствительных, двигательных, вегетативных — нервы могут быть полностью или частично смешанными и реже несмешанными. Передние ветви спинальных нервов формируют соматические сплетения (шейное, плечевое, поясничное и крестцово-копчиковое) и межреберные нервы. Вегетативные нервы формируют много вне- и внутриорганных сплетений.
Межпозвоночные (спинномозговые) узлы — 31 пара располагаются по выходе из позвоночного канала в межпозвоночных отверстиях. Они содержат ложно униполярные клетки, которые своими короткими отростками формируют задние, чувствительные корешки, а длинными входят в состав спинномозговых нервов. Такие же клетки находятся в чувствительных узлах черепных нервов: тройничного, лицевого и промежуточного, языкоглоточного, блуждающего.
Простая рефлекторная дуга состоит из двух нейронов: афферентного и эфферентного. В ней импульс продвигается от чувствительного нервного окончания, принадлежащего аксону афферентного нейрона к его телу, где переходит на дендриты, которые контактируют с эфферентным нейроном и по его аксону достигает органа. Такая дуга характерна для вегетативной системы.
Сложная рефлекторная дуга включает цепочку нейронов от трех и более. В ней между афферентным (рецепторным) и эфферентным (исполнительным) нейронами располагается один, а чаще несколько ассоциативных нейроцитов. Таких дуг больше встречается в соматической системе.
На основе построения рефлекторных дуг отечественные ученые И. М. Сеченов, С. П. Боткин, И. П. Павлов, П. К. Анохин, Н. М. Бехтерева разработали и усовершенствовали теорию нервизма. И. М. Сеченов предложил идею причинности (детерминизма), по которой считал, что всякое явление в организме имеет причину и в ответ на ее воздействие возникает рефлекторная реакция. И. П. Павлов доказал, что все виды деятельности обусловлены рефлексами. Простые, врожденные, видовые формы жизнедеятельности (инстинкты) возникают на основе безусловных рефлексов. Сложные формы, социально-интеллектуальные, работают на основе условных рефлексов. Благодаря рефлексам формируется первая и вторая сигнальные системы, деятельность которых обеспечивается рефлекторными дугами анализаторов. П. К. Анохин установил обратную связь любого органа с нервными центрами. Н. М. Бехтерева открыла в 80-ые годы прошлого века особые «смысловые» нейроны, осуществляющие аналитический выбор и передачу информации.
Билет 34
Поясничный и крестцовый отделы симпатического ствола, его топография, узлы, ветви, области, иннервируеимые ими.
Поясничный отдела симпатического ствола представлен чаще 3-5 (от 2 до 7) поясничными узлами и соединяющими их межузловыми ветвями.
Поясничные узлы веретенообразные, размеры не превышают 6 мм. Они располагаются на переднебоковой пов-сти тел поясничных позвонков и покрыты внутриюрюшной фасцией. К поясничным узлам правого симпатического ствола спереди прилежит нижняя полая вена. Узлы левого ствола примыкают к брюшной части аорты. Поясничные узлы этих симпатических стволов соединятся ветвями.
от каждого поясничного узла отходят 2 вида ветвей:
1) Серые соединительные ветви (содержат постганглионарные симпатические волокна, идущие к поясничным спинномозговым нервам)
2) Поясничные внутренностные нервы (имеют как преганглионарные, так и постганглионарные симпатические волокна, идущие к чревному сплетению и сосудистым вегетативным сплетениям)
Крестцовый отдел симпатического ствола образован 4 крестцовыми веретенообразными узлами, соединенными межузловыми ветвями.
Крестцовые узлы лежат на тазовой пов-сти крестца. Внизу правый и левый симпатические стволы сближаются и заканчиваются общим непарным узлом. Как и в поясничном отделе, между узлами стволом правой и левой сторон имеются соединения.
От крестцовых узлов отходит 3 вида ветвей:
1) Серые соединительные ветви (в из составе послеузловые симпатические волокна, идущие к крестцовым и копчиковому спинномозговым нервам и далее - в области, где они разветвляются)
2) Крестцовые внутреностные нервы (идут к верхнему и нижнемц чревным вегетативным сплетениям)
3) Органные ветви (идут к органным сосудистым сплетениям малого таза).
Билет 35
Преддверно-улитковый нерв, его анатомия, топография, области иннервации.
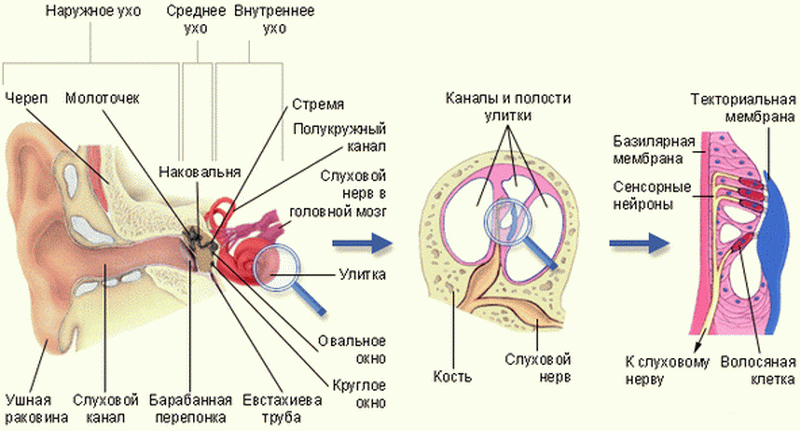
Преддверно-улитковый нерв (vestibulocochlearis) образован чувствительными волокнами, идущими от органов слуха и равновесия. На вентральной пов-сти ГМ нерв выходит позади моста, латерально от лицевого нерва. Далее он идет во внутренний слуховой проход, где разделяется на преддверную и улитковую части.
1) Преддверная часть образована отростками биполярных нейронов преддверного узла, расположенного на дне внутреннего слухового прохода. Периферические отростки нейронов преддверного узла образуют: передний, задний и латеральный ампулярные нервы; сферически-мешотчатый нерв. Все эти тонкие нервы заканчиваются рецепторами в перепончатом лабиринте внутреннего уха. Центральные отростки этих нейронов образуют преддверную часть преддверно-улиткового нерва, идущую в преддверным ядрам ствола ГМ.
2) Улитковая часть образована центральными отростками биполярных нейронов улиткового узла - спиральногоузла улитки, находящегося в спиральном канале улитки. Центральные отростки жтих нейронов идут к расположенным в покрышке моста улитковым ядрам. Периферические отростки начинаются в спиральном органе улитки внутреннего уха.
Волокна улитковой части оканчиваются в дорсальном и вентральном ядрах покрышки моста, а волокна преддверной части - в ядрах, расположенных в ромбовидной ямке. Значительная часть волокон преддверной части направляется в задний продольный пучок, вестибулярный спинальный пучок, а также в мозжечок. Волокна улитковой части частично перекрещиваются в составе боковой петли к нижним бугоркам четверохолмия и к медиальному коленчатому телу (отсюда начинается центральный слуховой путь).
Билет 36
Преломляющие среды глазного яблока (роговица, жидкость камер глаза, хрусталик, стекловидное тело, их анатомическая характеристика)
Роглвица (cornea) лишена сосудов.Она имеет вид часового стекла, выпуклого спереди и вогнутого сзади. Диаметр - 12 мм, толщина - 1 мм. Периферический край - лимб роговицы как бы вставлен в передний отдел склеры, в которвую переходит роговица. У роговицы различают 5 слоев:
1) Передний эпителий: представлен 5-6 слоями эпителиальных клеток. Этот многослойный плоский неороговевающий эпителий содержит многочисленные нервные окончания, обладает высокой регенеративной способностью)
2) Передняя пограничная пластинка расположена под базальной мемьраной, имеет фибриллярное строение.
3) Собственное вещество роговицы представлено распологающимися под углом друг к другу соединительно-тканными пластинками, они образованы пучками коллагеноых волокон. Между этими волокнами расположены клетки. Эти клетки и плстинки находятся в аморфном в-ве.
4) Задняя пограничная пластинка образована коллагеновыми волокнами, также залегающими в аморфном в-ве
5) Зданий эпителий роговицы образован плоскими клетками полигональной формы.
Так как роговица не имеет кровеносных сосудов, все обменные процессы происходят через внутриглазную и слезную жидкость, а также через сосуды, расположенные вокруг нее. Иннервация осуществляется чувствительнвми, вегетативными и трофическими нервами. Роговица выполняет защитную и опорную ф-и.
Водянистая влага камер глаза - прозрачная жидкость, заполняющая переднюю и заднюю камеры глаза. По своему составу она похожа на плузму крови, но имеет меньшее содержание белка. Она образуется специальными непигментированными клетками из крови. Водянистая влага содержит питательные в-ва(аминоксилоты, глюкоза). Благодаря присутсвию в ней иммуноглобулинов и своей постоянной циркуляции она способствует удалению потенциально опасных фактором из внутренней части глаза, а также обуславливает внутриглизаное давление.
Стекловидное тело, покрытое по периферии мембраной, находится в стекловидной камере, позади хрусталика, где плотно прилежит к внутренней пов-сти сетчатки. Хрусталик как бы вдавлен в переднюю часть стекловидного тела, которое в этом месте имеет углубление - стекловидную ямку. Стекловидное тело представляет собой желеобразную массу, лишенную сосудов и нервов.
Хрусталик располагается позади камер глаза, имеет форму двояковыпуклой линзы, обладающей большой светопрлеломляющей способностью. Передняя пов-сть хрусталика и наиболее выступающая ее точка - передний полюс, обращены в сторону задней камеры глаза. Более выпуклая задняя пов-сть и задний полюс прилежат к передней пов-сти стекловидного тела. Условная линия, соединяющая полюсы, назывется осью хрусталика (она совпадает с оптической осью глаза). Закругленный периферический край, где сходятся передняя и задняя его пов-сти - экватор. Вещество хрусталика бесцветное, плотное, прозрачное, сосудов и нервов не содержит. Внутренняя часть - ядро хрусталика, значительно плотнее, чем периферическая - кора хрусталика. Снаружи хрусталик покрыт тонкой прозрачной эластической капсулой. Способность хрусталика к видению на различные расстояния - аккомодация. Поэтому хрусталик вместе с ресничной мышцей и соединяющими их волокнами называют аккомодационным аппаратом глаза.

Билет 37
Проводящие пути проприоцептивной чувствительности: их положение в различных отделах спинного и головного мозга.
Пути проприоцептивной чувствительности идут к коре полушарий и в мозжечок. Проводящий путь проприоцептивной чувствительности в СМ проходит в заднем канатике, где образует тонкий пучок (Голля) - передают импульсы от нижних конечностей и нижней половины тела, и латерально располоденный клиновидный пучок (Бурдаха) - несет импульсы от верхних конечностей и верхней половины тела.
Первые нейроны располагаются в чувствительных узлах спинномозговых нервов, а вторые - в продолговатом мозге. Отростки вторых нейронов в продолговатом мозге образуют перекрест мудиальных петель, а затем, в пределах ствола ГМ формируют бульботаламический путь (медиальную петлю). Часть волокон 2 нейрона повыходе из тонкого и клиновидного ядер сгибаются кнаружи и образуют наружные дорсальные и вентральные дугообразные волокна, которые через нижние мозжечковые ножки идут к коре червя мозжечка. Медиальная петля проходит в покрышке моста и среднего мозга, ее волокна заканчиваются в таламусе (3-й нейрон), отростки 3-х нейронов проходят в задней ножке внутренней капсулы и идут в кору большого мозга (постцентральную извилину).
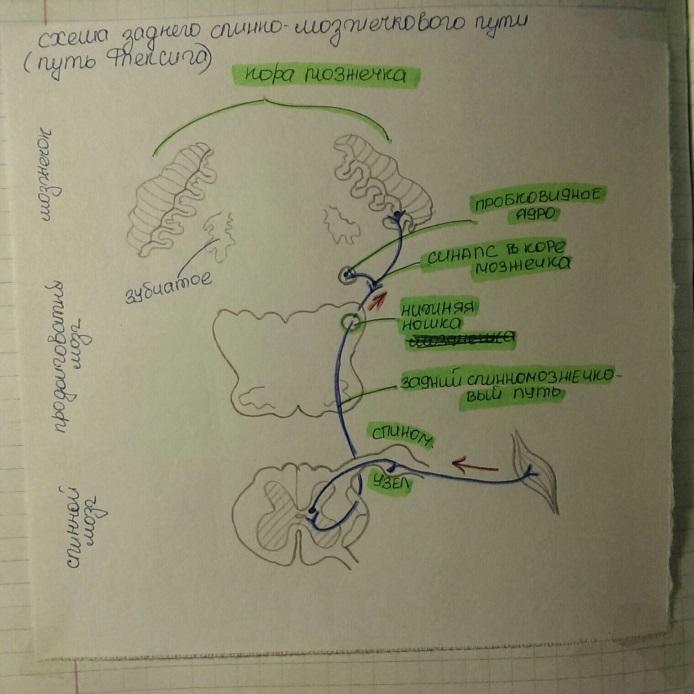
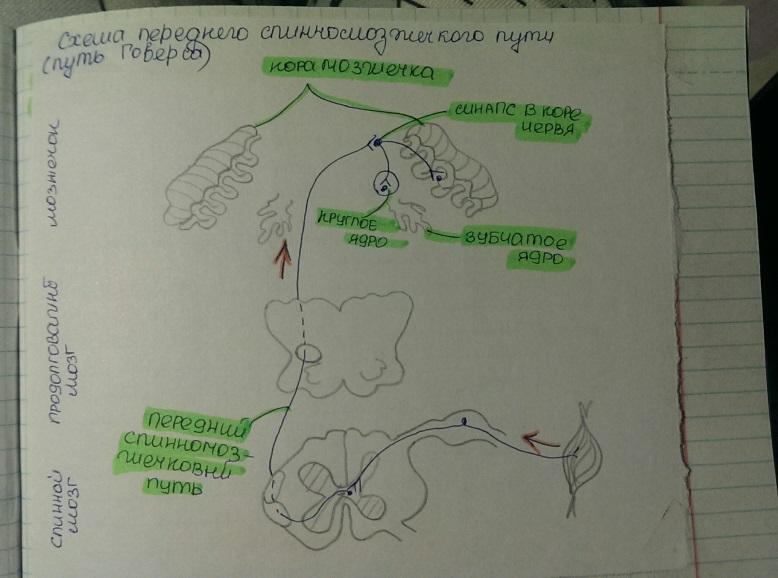
Проприоцептивные пути, идущие к мозжечку, передают информацию о состоянии опорно- двигательного аппарата. Они представлены задним (неперекрещенным) и передним (дважды перекрещенным) спинномозжечковыми путями.
Задний спинномозжечковый путь передает импульсы от мышц, сухожилий, суставов в мозжечок. Тела клеток 1-го нейрона находятся в спинномозговом узле, а их центральны отростки в составе заднего корешка идут в задний рог СМ и заканчиваются синапсами на клетках грудного ядра. Клетки грудного ядра явлеются 2-м нейроном. Аксоны этих клеток выходят в боковой канатик своей стороны, в его заднюю часть, поднимаются вверх и через нижнюю мозжечковую ножку входят в мозжечок, к клеткам коры червя.
Передний спинномозжечковый путь. Тело клетки 1-го нейрона располагается в спинномозговом узле. Его периферический отросток имеет окончания в мышцах, сухожилиях, суставных капсулах. Центральный отросток 1-го нейрона в составе заднего корешка входит в СМ и заканчивается синапсами на клетках, примыкающих к грудному ядру. Аксоны клеток этого 2-го нейрона проходят через переднюю серую спайку в боковой канатик противоположной червя стороны, в его переднюю часть. Т.о. передний спинномозжечковый путь проделывает дважды перекрещенный путь, возвращается на ту же сторону, на которой возникли проприоцептивные импульсы. Проприоцептивные импульсы, поступившие в кору червя, также передаются в красные ядра и через зубчатое ядро к коре ГМ.
Схема проведения пути проприоцептивной чувствительности коркового направления

Передний спинномозжечковый путь
Задний спинномозжечковый путь