

21. Пути ассимиляции со2
Цикл Кальвина: сост-т из 3-х этапов:1)карбоксилирование; 2)восстановление; 3)регенерация акцептора СО2.
1) Карбоксилир-е: первичный акцептор СО2 - это 1,5-РДФ (рибулозо-1,5-дифосфат), а ключевой фермент – РДФ-карбоксилаза. На 1-ой стадии образ-ся нестойкое 6-тиуглеродное соед-е, кот-е быстро распад-ся на 2-е триозы – 3ФГК (3-фосфоглицериновая к-та). 2)Восстан-е: восстан-е 3ФГК до 3ФГА происходит в 2 этапа. Вначале при участии АТФ и фосфоглицераткиназы 3ФГК присоед-т остаток Н3РО4 с образованием 1,3-диФГК. Это повыш-т реакционную способ-ть соед-я и оно восстанавл-ся с помощью НАДФ-Н до 3ФГА. 3) Регенер-я: в рез-те 2-х 1-ых стадий при фиксации 3-х мол-л СО2 образов-сь 6 мол-л восстановленных фосфотриоз, пять из них использ-ся для регенерации РДФ, а одна- для синтеза глюкозы. (т.о. для образ-я 1-ой мол-лы глюкозы цикл должен пройти 2 раза).
Цикл Хэтча-Слэка-Карпилова (С4-путь): С4-раст-я – это растения жаркого климата, кот-е осущ-т фиксацию СО2 при закрытых устьицах. В листья у них содер-ся хлоропласт 2-х типов:1) в кл-х мезофилла – хлороп-т обычного типа; 2) в кл-х обкладки – агранальные хлороп-ты(без гран). Акцептор СО2 – ФЕП(фосфоенолпируват), а основной фермент – ФЕП-карбоксилаза. В кл-х мезофилла: СО2 вступает в реакцию с ФЕП и образ-ся ЩУК, кот-я с помощью НАДФ-Н восстан-ся до яблочной к-ты.Яблочная к-та переносится в хлороп-ты кл-т обкладки. В Кл-х обкладки: яблочная к-та подверг-ся окислительному декарбоксилированию и образ-ся ПВК, кот-я снова переходит в кл-ки мезофилла, где при участи АТФ регенерирует в ФЕП. Образовавшиеся в рез-те окислит-го декарбоксилирования СО2 и НАДФ-Н поступают в цикл Кальвина, учавствуя в синтезе сахаров.. Т.О. в данном случае процесс карбоксилирования и образования сахаров разделен в пространстве.
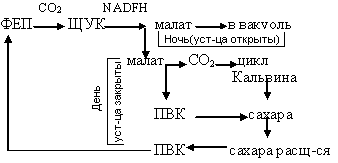
САМ-тип(С4-раст.): метаболизм по типу толстяноквых – осущ-т суккуленты, для кот-х харак-но разделения прцессов карбоксилирования и образ-я сахаров не в пространстве, а во времени. Ночью (уст-ца открыты): СО2 из атмос-ры при участии ФЕП-карбоксилазы реагирует с ФЕП и обр-ся ЩУК, кот-я восстан-ся до малата и накаплив-ся в вакуолях(подкисляется клеточный сок). Днем (уст-ца закрыты): образовав-ся малат транспортир-ся в цитоплазму, декарбоксилир-ся с образ-м ПВК (учавствует с с/зе сахаров) и СО2, кот-ый вкл-ся в цикл Кальвина и учавст-т в синтезе сахаров.
22.Строение этц дыхания растений. Субстратное и окислительное фосфорилирование.Альтернативный механизм переноса электронов.
Дыхательная ЭТЦ митохондрий локализована во внутр. мембране и служит для передачи Н+ и е- от восстанавливаемых субстратов на мол-лы О2. ЭТЦ растений сос-т из 4-х мультибелковых комп-в и 2-х небольших компонентов Q(убихинон) и С(цитохрома с).
I) Содержит: ФМН(флавинмононуклеоитд)-кофермент дегидрогеназы, окисляющей эндогенный НАД-Н и FeS-железосерные белки. Копл-с I осущ-т перенос Н+ и е- от НАД-Н к Q. Субстрат для комп-са I- НАД-Н из ЦТК. II) Сод-т: ФАД(флавинадениндинуклеотид) – кофермент сукцинатдегидро-зы и железосерные белки. Функ-я – осущес-т окисление сукцината(янтарная кислота) убихиноном. III) Состав: цитохромы b и c, FeSR(железосерный белок Риске). Функ-я -переносит Н+ и е- от восстановл-го Q к цитохрому С, по строению III сходен с цитохромным комплексом тилакоидов хлоропластов. IV) Состав: цитохрома а и а3 , функционирующие в комплексе с атомами Сu. Терминальный комп-с, переносит Н+ и е- с цитохрома С на молекул-ый О2. Комлексы I , III и IV явл-ся пунктами сопряжения транспорта е- и окислит-го фосфорилир-я(т.е. там образ-ся АТФ из АДФ и неорганич-го фосфата). Кроме переноса е--в по ЭТЦ имеет место также трансмембранный перенос Н+ и формир-е ΔμН (трансмембран-го электрохим-го потенциала) т.е. ЭТЦ митохондрий выполн-т роль протонной помпы: «+»заряды скаплив-ся на внешн.поверх-ти мембраны, «-»заряды на внутрен. поверхности – образ-ся ΔμН; Е запасенная в виде ΔμН, быстро превращ-ся в более стабильную форму – Е макроэргических связей в мол-ле АТФ. Образов-е АТФ из АДФ и неоган-го фосфата и есть суть процесса окислительного фосфорилиров-я.
Т.О. образ-е макроэргических соед-ий в растит. кл-ке м. свести к 6-ти реакциям(3 реакции субстратного фосфорилир-я и 3 реакции окислит-го фосфорил-я).
Субстратное фосфорилир-е:1) Окисл-е 1,3-диФГК (дифосфоглицериновая к-та) до 3ФГК в гликолизе; 2) Дегидратация 2ФГК через ФЕП(фосфоенлпируват) до енолПВК в гликолизе; 3) Окислительное декарбоксилирование ά-кетоглутаровой к-ты в ЦТК.
Основные черты субстратного фосфорилирования: протекает в раст-ре и не связано со структ-ми элем-ми кл-ки; донором явл-ся сам субстрат дыхания и макроэргич-кая связь образ-ся непоредст-но на окисляемом субстрате; в ходе окисления субстрат не регенерирует и не треб-ся в дальнейших реакциях; прочно сопряжено с синтезом АТФ.
Основные реакции окислит-го фосфорилир-я:1) Перенос Н+ и е- от НАД-Н к Q в комп-се I; 2) Перенос Н+ и е- от цитохр. b к цитохр.с черех комп-с III; 3) Перенос Н+ и е- от цитохром-го комп-са а-а3 к молек-му О2.
Альтернативное дыхание: в раст-х сущ-т альтернт-ый путь переноса е--в. В этом случае поток е--в обходит комплесы III и IV(на кот-х набл-ся сопряжение транспорта Н+ и е- и фосфорилиров-я ) и е--ны идут от убихинона прямо к молекулярному О2. Это возможно благодаря наличию в кл-х растений альтернативной терминальной оксидазы. Деятель-ть этого фермента не сопряжена с запасанием Е в виде АТФ, но альтернативная оксидаза обеспечивает меньшую чувств-ть растений к цианидам(по сравнению с жив-ми). Альтернативный путь окисления функц-т в услов-х высокого содерж-я АТФ и в случ-х, когдасниж-ся активность основной цепи дыхания, что способств-т окисл-ю НАД-Н. Альтернатив. дых. учасвуют в пр-ссе термогенеза: энергия, кот обр при окислении S дыхания расход на образование теплоты. Это обеспеч повышение t в тканях растения в период их цветения на 10-15 градусов.
23. Основные соединения серы врастении, их роль в структурной организации клетки, участие в ОВР. Сера явл.одним из основных элементов питания. Потребность в сере высокая у растений, богатых белками,например, у бобовых и крестоцветных, которые в большом количестве синтезирируют серосодержащие горчичные масла.Сера содержится в растениях в 2-х формах: окисленной,т.е в виде неорганического субстрата и в восстановленной. Сера поступает в корень в виде сульфата метаболизируясь входит в состав органических соединений (цистеина,цистина,КоА, тиамина и др.)Основная часть сульфата перемещается из корней в сосуды ксилемы и с током переносится молодым растущим органам, где она интенсивно включается в обмен и теряет подвижность. Редукция сульфата и синтез серосодержащих АК и белков наблюдается при созревании семян. Сера минимальна в молодых листьях и резко возрастает при их старении в связи с усилением процесса деградации серосодержащих белков. Фунции серы:1) участие SH-групп в образовании ковалентной и водородной связей поддерживающих 3-х мерную структуру белка. Дисульфидные мостики между полипептидными цепями или 2-мя участками одной цепи стабилизируют молекулу белка.2) Поддержание определенного уровня ОВ потенциала клетки, за счет необратимости реакций. 3) Сера входит в состав важнейших биологических соединений : КоА и витаминов, и в форме этих соединений принимает участие в энзиматических реакциях клетки. 4) Липолевая кислота явл.коэнзимом в растениях окиленного декарбоксилированием альфа-кетокислот. 5) Производные тиамина действуют как коэнзим при декарбоксилировании пирувата в ацетальдегиде и в окислении альфа-кетокислот. 6) Многие виды растений в малых количествах содержат летучие соединения серы, например, сульфооксиды входят в состав фитонцидов лука и чеснока, именно их присутствие вызывает раздражение глаз при нарезке лука. Недостаточное снабжение растений серой тормозит синтез серосодержащих АК и белков. Снижает фотосинтез и скорость роста растений(надземной части). В особых случаях нарушается формирование хлоропласта и возможен распад. Симптомы дефицита серы: побледнение и пожелтение листьев. Это свидетельствуют о том, что отток серы из более старых листьев не может компенсировать недостаточное снабжение растений серой через корни.
24.Гуттация, транспирация; физиологическое значение этих процессов,кол-ые показатели транспирации.Пр-ом раб. нижнего концевого двигателя служат т.н. «плач» растений и гуттация. Явл-ие «плача» свидет-ет о корневом давлении. Гуттация набл-ся при выс. влажности воздуха. В рез-те деят-ти нижнего концевого двигателя на концах и зубчиках листьев выд. капельно-жидкая влага — растение гуттирует. Характерно для листьев тропических растений, приспособленных к жизни в условиях повышенной влажности, при которых транспирация затруднена.
Транспирация — это физиологический процесс испарения воды растением. Осн-ым орг-м тран-ции явл-ся лист.Тран-ия слагается из двух процессов: а) передвиж. воды из листовых жилок в поверхностные слои стенок клеток мезофилла; б) испарения воды из кл-ых стенок в межклетные пространства и подустьичные полости с последующей диффузией в окружаю-щую атмосферу через устьица (устьичная транспирация) или испарения воды из клеточных стенок эпидермиса в атмосферу путем кутикулярной транспирации. Транспирация играет в жизни растения значительную роль: 1)она служит одним из ос-новных механизмом передвижения воды и различных веществ вверх по стеблю,2)оказывает большое влияние на процессы воздушного питания растения. Поглощение СО2 из воздуха осуществляется посредством устьиц, через которые одновременно испаряется основная масса воды, 3)защищает листья от перегрева. Только в условиях непрерывной транспирации температура листьев может поддерживаться на уровне, лишь незначительно превышающем температуру окружающего воздуха.4)Снижение интенсивности транспирации в результате недостатка воды существенно сдвигает обычный ход физиологических процессов в листе.
Кол-ные показатели тран-ии.1)интенсивность тран-ии выражают в граммах испаренной воды за 1 ч на единицу площади или на 1 г сухой массы;2)продуктивность транспирации — количеством граммов сухих веществ, образуемых при расходовании каждых 1000 г воды. Величиной, обратной продуктивности транспирации, является транспирационный коэффициент, т. е. число граммов воды, израсходованной при накоплении 1 г сухих веществ.Интенсивность транспирации у большинства растений составляет 15 —250 г м2 *ч днем и 1 — 20 г • м2* ч ночью.