

3. Первинна структура нуклеиновых кислот
Первинна структура нуклеиновых кислот — це послідовність розташування мононуклеотидов у полинуклеотидной ланцюги.
Структура стабілізується 3', 5'-фосфодиэфирными зв'язками. У загальному виді структура нуклеиновой кислоти виглядає в такий спосіб:
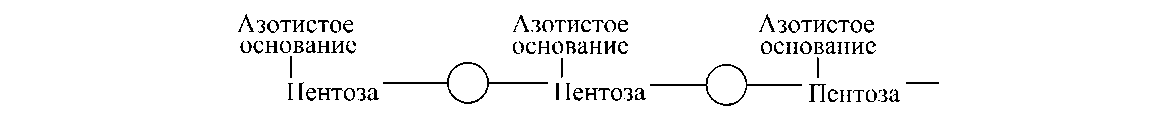
Межнуклеотидные зв'язки утворюють гідроксильні групи в 3'- і 5'-положеннях пентоз. Молекула РНК складається з однієї полинуклеотидной ланцюги, молекули більшості ДНК двухцепочечны.
Більшість клітинних РНК представлені одиночної полинуклеотидной ланцюжком: 5' цаааа—...уВ— —Ц— ц-ц-а—3'. На одному кінці РНК виявляється вільний монофосфорный ефір, цей кінець прийнято позначати як 5'-кінець. На протилежному кінці ланцюга (3'-кінці) розташовується нуклеотид з вільними 2'- і 3'-гідроксильними групами. трнк має рядом особливостей: 5'-кінцем її завжди є гуаниловая або цитидиловая кислота із залишком фосфорної кислоти, на 3'-кінці послідовність залишків мононукеотидов завжди постійна — ЦЦА. На частку мінорних нуклеотидов у структурі трнк доводиться 10%.
мрнк переносить генетичну інформацію від ДНК на белоксинтезирующий апарат клітки й визначає первинну структуру синтезованого білка. На 5'-кінці мрнк містить певну нуклеотидную послідовність, називану шапочкою (кэпом), на 3'-кінці — полиадениловая послідовність ( полиоа).
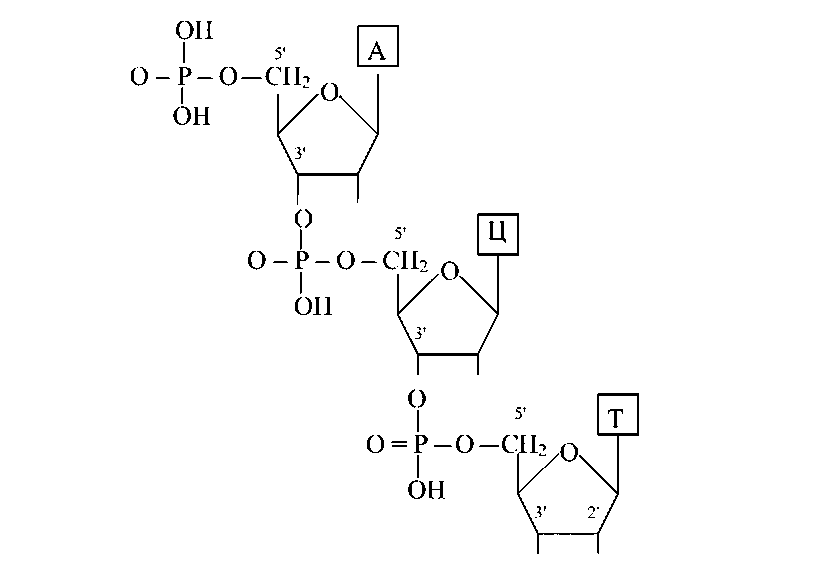
Первинна структура ДНК аналогічна такий у РНК, однак у силу великої молекулярної маси молекул ДНК визначення послідовності нуклеотидов утруднене. У цей час триває робота з розшифрування первинної структури гена людини.
4. Вторинна й третинна структура нуклеиновых кислот
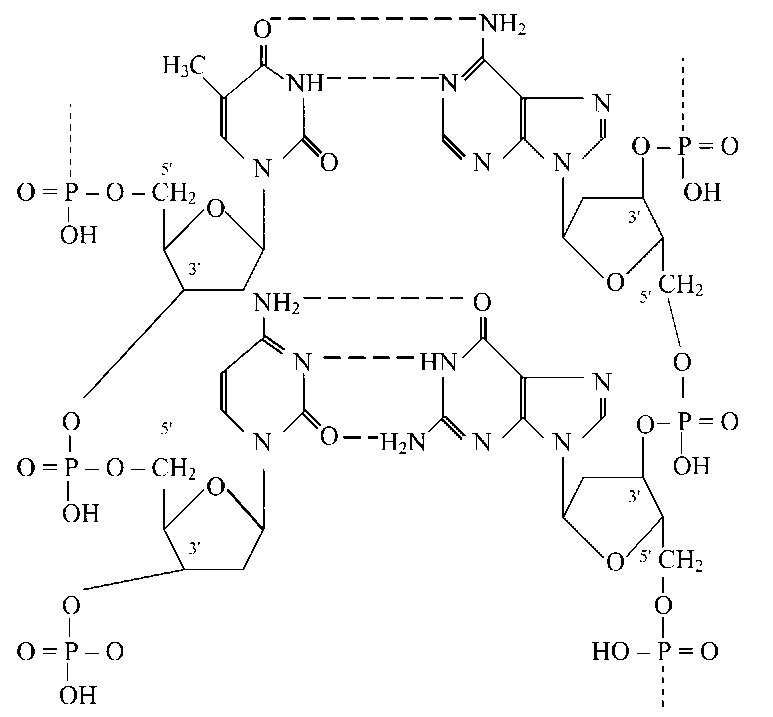
Вторинна структура нуклеиновых кислот — перший рівень компактизации нуклеотидной ланцюги. Установлене, що два ланцюги молекули ДНК утворюють правовращающуюся спіраль, у якій обидві ланцюги обертаються навколо однієї й тієї ж осі. Азотисті підстави в такій структурі перебувають усередині, а фосфорні й вуглеводні компоненти — зовні (модель УотсонанЛементу).
Азотисті підстави у двухспиральной молекулі ДНК покладені парами: пуриновій підставі з одного ланцюга відповідає пиримидиновое з інший, аденину відповідає тимин, а гуаніну — цитозин. Така виборча взаємодія називається комплементарним, а відповідні азотисті підстави — комплементарними. Стабільність пари А — Т забезпечується двома водневими зв'язками, а пари Г — Ц — трьома. Довжина зв'язків між підставами близько 0,3 нм.
Полинуклеотидные ланцюги в молекулі ДНК мають протилежну полярність: один ланцюг іде в напрямку 3'— 5', а інша — у напрямку 5'— 3'. Крок спирали молекули ДНК рівний 3,4 нм, діаметр двухспиральной молекули становить 1,8 нм.
Залежно від кількісного змісту води й іонної сили навколишнього середовища конфігурація подвійної спіралі ДНК може мінятися. Доведене існування шести форм ДНК, названих A, B, C, D, E і ZмФормами. В AмФорми відзначається зсув пар підстав від центру до периферії, крок спирали в цієї форми менше (2,8 нм), а діаметр спирали й кількість нуклеотидов на витку більше, чим в BмФорми класичною формою, що вважається, молекули ДНК. У структурі A- і BрФорм паралельно фосфодиэфирному кістяку йдуть мала й більша борозенки — місця приєднання білків. Z-Форма має левозакрученную конфігурацію, фосфодиэфирный кістяк розташований зигзагообразно уздовж осі молекули. Вторинна структура матричних і рибосомных РНК вивчена незначно. Передбачається, що вторинна структура т-РНК являє собою утвір на зразок конюшинового аркуша. Укладання РНК у вторинну структуру завжди йде відповідно до програми ДНК.
Третинна структура нуклеиновых кислот — другий рівень компактизации нуклеиновых кислот. Подвійна спіраль ДНК на деяких ділянках може зазнати суперспирализации з утвором суперспіралі або кільцевої форми. Утвір супервитків катализируется специфічними ферментами — топоизомеразами. Суперспирали з'єднуються з білкамиигистонами, що розташовуються в борозенках, цим забезпечується стабільність третинної структури ДНК.
Існує третинна структура трнк, яка має більшу компактність у порівнянні із вторинною структурою. Існують природні двухцепочечные РНК, що володіють подібної із Днк-структурою (у реовирусов).
У стабілізації третинної структури нуклеиновых кислот велике значення мають вандерваальсовы сили.