

Репродукция вирусов
Вирусы - облигатные внутриклеточные паразиты, способные размножаться только в живой клетке. Для того чтобы вирус мог размножиться, он должен вначале заразить клетку. Спектр хозяев определяется как типами клеток, так и видами животных, которых он может заражать и в которых он способен размножаться. Спектр хозяев разных вирусов значительно варьирует. Одни вирусы имеют широкий спектр хозяев, другие заражают лишь клетки одного типа определенных видов животных'. Способность клетки или животного заражаться называют восприимчивостью, а способность обеспечить репродукцию вируса - пермиссивностью. Восприимчивость клеток не следует путать с пермиссивностью; так, клетки кур не восприимчивы к вирусу полиомиелита, поскольку у них нет рецепторов для связывания с вирионами. Однако они полностью пермиссивны в отношении этого вируса, так как они продуцируют вирус после заражения их интактной РНК, экстрагированной из вирусных частиц.
В отличие от прокариотических и эукариотических микроорганизмов вирусы не размножаются бинарным делением. Размножение вирусов происходит путём репродукции (англ, "reproduce" - воспроизво-аить, делать копию), то есть воспроизведение их нуклеиновых кислот и белков c последующей сборкой вирионов. Синтез нуклеиновых кислот и белков вируса происходит в разных частях клетки (ядре и цитоплазме). Такой способ репродукции получил название дизъюнктивного (разобщённого).
Цикл размножения
Большое влияние на изучение размножения вирусов животных оказали более ранние работы по бактериальным вирусам; в частности, понятие «единичного цикла размножения» возникло при исследовании цитоцидных инфекций, вызванных вирулентными бактериофагами. Эллис и Дельбрюк (1939) разработали постановку эксперимента с одноступенчатым размножением путем подбора таких условий, при которых поведение популяции зараженных клеток в целом могло быть использовано для проверки предположений о том, что происходит в любой индивидуальной клетке в данный момент времени после начала инфекции. По существу «одноступенчатый» опыт в том виде, в каком он применяется для вирусов животных, состоит в одновременном заражении популяции клеток с множественностью не менее одной инфекционной вирусной частицы на клетку. Для того чтобы обеспечить заражение каждой клетки и высокую степень синхронности последующих событий в популяции зараженных клеток, множественность заражения должна составлять 10—100 частиц. После инактивации неадсорбированного вирусного материала через небольшие интервалы времени в культуре определяют число образовавшихся инфекционных вирусных частиц, как внутриклеточных, так и вышедших в среду (рис. 5-1).
Термин эклипс в настоящее время представляет исторический интерес; он часто фигурировал в дискуссиях о размножении вирусов 10 и более лет назад и отражает тот факт, что заражающий вирион исчезает и его нельзя обнаружить в клетке на протяжении нескольких часов периода эклипса до того момента, когда появляются частицы вирусного потомства. Термин латентный период относится к интервалу между исчезновением внесенных вирионов и выходом в среду новых вирусных частиц. В отличие от бактериофагов для вирусов животных характерен постепенный выход вирионов из клетки поодиночке на протяжении длительного периода, причем иногда большая часть вирионов остается связанной с клеткой в течение всего периода размножения вируса. Даже если вирусы разрушают плазматическую мембрану, клетки животных не «взрываются», как бактерии, а несколько сморщиваются, и основная часть вирусного потомства может оставаться заключенной в клеточном детрите.
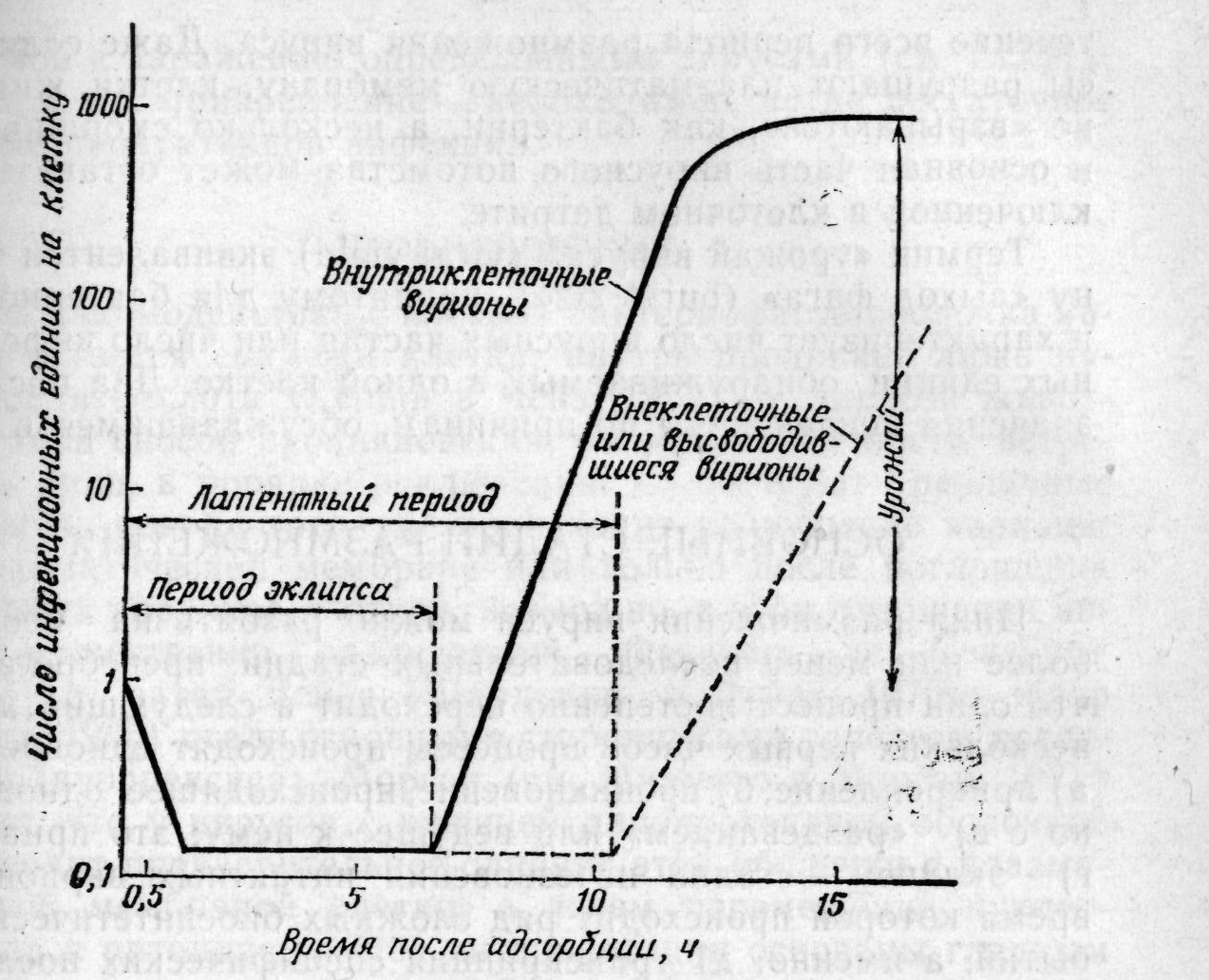
Схема цикла размножения вирусов животных.
Вирусы выходят из клетки на поздней стадии и не полностью. В других случаях,
когда вирионы созревают в процессе выхода из клетки, кривые для внутриклеточной
и внеклеточной инфекционности совпадают.
Чтобы заразить клетку, вирион должен связаться с клеточной поверхностью, проникнуть в клетку и «раздеться» до такой степени, чтобы его геном стал доступен для вирусного или хозяйского аппарата, обеспечивающего транскрипцию или трансляцию.
Процесс репродукции вирусов условно можно разделить на 2 фазы. Первая фаза включает 3 стадии: 1) адсорбцию вируса на чувствительных клетках; 2) проникновение вируса в клетку; 3) депротеинизацию (раздевание вируса) вируса. Вторая фаза включает стадии реализации вирусного генома: 1) транскрипцию, 2) трансляцию, 3) репликацию, 4) сборку, созревание вирусных частиц и 5) выход вируса из клетки.
Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. с прикрепления вируса к поверхности клетки.
АДСОРБЦИЯ (прикркпление).
Адсорбция представляет собой специфическое связывание вирионного белка (антирецептора) с комплементарной структурой клеточной поверхности - клеточным рецептором. Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусных частиц к клеточной поверхности. Процесс адсорбции возможен при наличии соответствующих рецепторов на поверхности клетки и «узнающих» их субстанций на поверхности вируса. Антирецепторы распределены по поверхности вирусов, заражающих клетки человека и животных. Сложные вирусы, такие как вирус осповакцины (поксвирус) и вирус простого герпеса (герпесвирус), могут иметь антирецепторные молекулы нескольких видов. Более того, молекулы антирецепторов могут иметь несколько доменов, каждый из которых взаимодействует с определенным рецептором. Чувствительность клеток ограничена доступностью рецепторов, и не все клетки в восприимчивом организме экспрессируют рецепторы. Почечные клетки человека в структуре почек не имеют рецепторов для вируса полиомиелита, но при культивировании этих клеток in vitro рецепторы появляются. Мутации генов, кодирующих антирецепторы, иногда ведут к потере их способности взаимодействовать с рецепторами.
Самые начальные процессы адсорбции имеют неспецифический характер, и в основе их может лежать электростатическое взаимодействие положительно и отрицательно заряженных группировок на поверхности вируса и клетки. Адсорбция происходит лишь при наличии определённых электролитов, в частности ионов Са2+, которые нейтрализуют избыточные анионные заряды вируса и клеточной поверхности и уменьшают электростатическое отталкивание. Адсорбция вирусов мало зависит от температуры и энергии.
Затем наступает специфическое взаимодействие прикрепительного белка вириона со специфическими группировками на плазматической мембране клетки. Простые вирусы человека и животных содержат прикрепительные белки в составе капсида. У сложно организованных вирусов прикрепительные белки входят в состав суперкапсида. Они могут иметь форму нитей (фибры у аденовирусов), либо шипов, грибоподобных структур у миксо-, ретро-, рабдо- и других вирусов. Однако узнавание клеточных рецепторов вирусными белками, ведущее к прикреплению вирусной частицы к клетке, является высоко специфическим процессом. Вирусы используют рецепторы, предназначенные для прохождения в клетку необходимых для ее жизнедеятель ности веществ: питательных веществ, гормонов, факторов роста и т. д. Рецепторы могут иметь разную химическую природу и представлять собой белки, углеводный компонент белков и липидов, липиды.
По химической природе рецепторы, на которых фиксируются вирусы, относятся к двум группам: муко(глико)протеидным и липопротеидным. Вирусы гриппа, парамиксовирусов (парагриппа), аденовирусы фиксируются на мукопротеидных рецепторах (сиаловая кислота в составе гликопротеидов и гликолипидов - ганглиозидов). Энтеровирусы, вирусы герпеса, арбовирусы адсорбируются на липопротеидных рецепторах клетки. Для рабдовирусов и реовирусов — также углеводный компонент в составе белков и липидов, для пикорна- и аденовирусов — белки, для некоторых вирусов — липиды. Специфические рецепторы играют роль не только в прикреплении вирусной частицы к клеточной поверхности. Они определяют дальнейшую судьбу вирусной частицы, ее внутриклеточный транспорт и доставку в определенные участки цитоплазмы и ядра, где вирус способен инициировать инфекционный процесс. Вирус может прикрепиться и к неспецифическим рецепторам и даже проникнуть в клетку, однако только прикрепление к специфическому рецептору приведет к возникновению инфекции. Вначале происходит единичная связь вириона с рецептором - такое прикрепление непрочное - адсорбция носит обратимый характер.
Для того чтобы наступила необратимая адсорбция, должны появиться множественные связи между вирусной частицей и многими молекулами рецепторов, т. е. должно произойти стабильное мультивалентное прикрепление. Количество молекул клеточных рецепторов в участках адсорбции может доходить до 3000. Количество специфических рецепторов на поверхности одной клетки составляет 10³-104. Рецепторы для некоторых вирусов, например, для арбовирусов, содержатся на клетках как позвоночных, так и беспозвоночных, для других вирусов - только на клетках одного или нескольких видов.