

2.6. Каріотип
Сукупність набору хромосом клітини і, загалом, організму називають каріотипом. Каріотипування відіграє важливу роль у в ідентифікації хромосом. Для цього набір хромосом мета фазної клітини фотографують або змальовують за допомогою рисувального апарату. Після цього в результаті довгого ототожнення гомологічних хромосом складають каріограми організму.
Індивідуальна характеристика хромосом в каріотипі показує, що хромосомні набори індивідуальних видів різноманітні і відрізняються не лише по формі, а й по розміру. Аналіз каріотипів багатьох видів показав, що не існує в природі двох однакових каротипів, азначить і видів.
3. Молекулярні основи спадковості
3.1.Особливості будови днк
Розшифрування структури молекули ДНК, виконане американським вірусологом Дж. Уотсоном та англійським фізіологом Ф. Кріком в 1953р. допомогло їм створити молекулярну модель цього складного полімеру (рис. 6). Головні риси моделі були описані Уотсоном і Кріком, коли вони дійшли висновку, що ДНК – це високомолекулярний полімер, який складається з двох полі нуклеїнових ланцюгів [2].
Р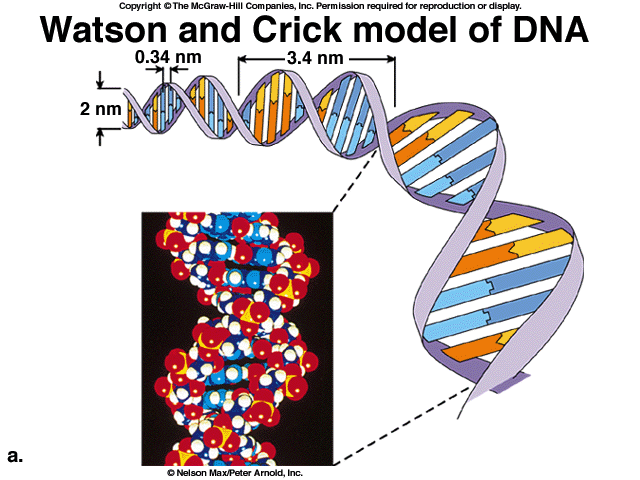 ис.
6. Модель організації ДНК Уотсона-Кріка.
ис.
6. Модель організації ДНК Уотсона-Кріка.
Мономерами кожного ланцюга є дезоксирибонуклеотиди. Розмір кожного нуклеотиду в поздовжньому вимірі молекули дорівнює 3.4 А0.
Ці ланцюги утворюють право закручені спіралі, кожний виток якої містить по 10 основ. Ланцюги полімеру закручені в спіраль один навколо другого і навколо загальної осі. Фосфатні групи цих ланцюгів містяться із зовнішньої сторони, а азотисті основи – всередині спіралі.
Довжина одного витка спіралі дорівнює 32 А0, а її діаметр – 20 А0. Ланцюги спіралі утримуються між собою за допомогою водневих зв’язків, які утворюються між парами аденін – тимінового (А - Т) та гуанін – цитозинового (Г- Ц) дизоксорибонуклеотидів. Пари А – Т та Г – Ц є специфічними в складі подвійної спіралі бо в нормі саме між гуаніном та цитозіном та між аденіном і тиміном у складі спіралі утворюються водневі зв’язки.
М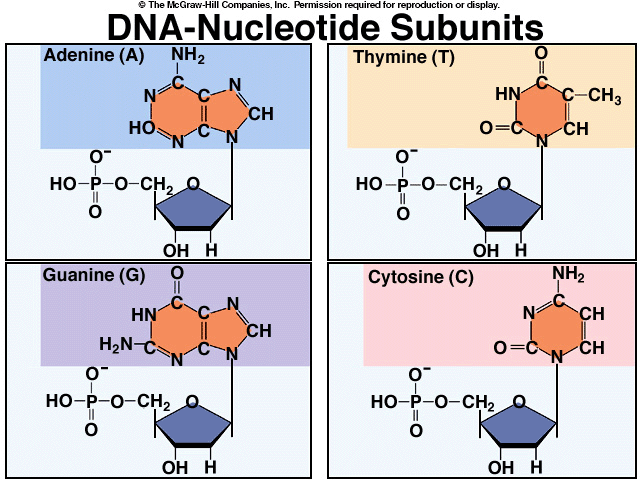 ономером
молекули ДНК є дезоксирибонуклеотид,
до складу якого входять залишок молекули
фосфорної кислоти, цукор дизоксорибоза
та одна з чотирьох азотистих основ. Ці
основи у складі ДНК представлені пуринами
(аденін та гуанін) і піримідинами (тимін
та цитозин) (рис. 7).
ономером
молекули ДНК є дезоксирибонуклеотид,
до складу якого входять залишок молекули
фосфорної кислоти, цукор дизоксорибоза
та одна з чотирьох азотистих основ. Ці
основи у складі ДНК представлені пуринами
(аденін та гуанін) і піримідинами (тимін
та цитозин) (рис. 7).
Рис. 7. Нуклеотиди ДНК.
Всі типи нуклеотидів: аденіновий, гуаніновий, цитозиновий та тиміновий чергуються будови ланцюга в найрізноманітніших послідовностях. Послідовність головних валентностей нитки ДНК утворюються ефірними зв’язками між фосфорною кислотою і дезоксирибозоюю. Ці зв’язки утворюються атомами вуглецю молекулами цукру в 3’ та 5’ положенні.
Одна з важливих особливостей ДНК описується правилом еквівалентності, сутність якого полягає в тому, що в молекулах дезоксирибонуклеїнових кислот молярні відношення пуринів до піримідинів (або навпаки) дорівнюють одиниці (рис. 8). Ця закономірність відкрита Е.Чаргафом в 1950р. відіграла важливу роль у створенні дволанцюгової моделі молекули ДНК [3].
Р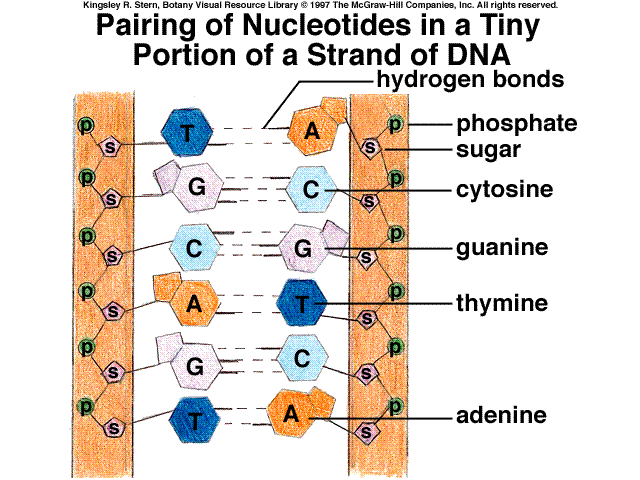 ис.
8. Комплементарність нуклеотидів в
подвійному ланцюгу ДНК.
ис.
8. Комплементарність нуклеотидів в
подвійному ланцюгу ДНК.
Спираючись на створену модель, Уотсон і Крік дійшли висновку, що оскільки будова молекули полімеру монотонно повторюється цукрово-фосфатні послідовності, а мінливими виявляються тільки послідовності азотистих основ, що генетична інформація може бути закодованою лише за допомогою послідовностей азотистих основ, які входять до складу нуклеотидів у молекулах ДНК можуть спричиняти лише мутації. Отже, була виявлена специфічність взаємного розташування азотистих основ, яку почали розглядати як матеріальну основу генетичного коду.