

Модулирующие нейроны
В нервной системе выделена особая группа клеток — модулирующих нейронов, которые сами не вызывают реакции, но регулируют активность других нейронов. Они образуют контакты с другими нейронами типа «синапс на синапсе». Модулирующие нейроны причастны к регуляции болевой чувствительности. Схема взаимодействия модулирующего нейрона с сенсорным, проводящим сигналы о болевом раздражении, описана Л. Иверсон (1984) следующим образом. Синаптическая передача болевых сигналов в головной мозг, опосредуемая веществом Р, находится под контролем нейропептидов, подавляющих болевые сигналы. В задних рогах спинного мозга вставочные модулирующие нейроны, выделяющие нейропептид—энкефалин, образуют синапсы на аксонных окончаниях болевых нейронов. Энкефалин тормозит выход вещества Р, что уменьшает возбуждение постсинаптического нейрона) посылающего в головной мозг сигналы о боли.
Функции модулирующих нейронов наиболее хорошо изучены на моллюсках. Показана причастность модулирующих серотонинергических нейронов виноградной улитки к регуляции состояния организма. Улитку, которая ползала по шару, плавающему в воде, в течение 8 дней подвергали сильному электрическому раздражению вне сочетания с тестирующим стимулом — слабым тактильным раздражением. Это увеличивало величину реакции улитки на тактильный стимул — втягивание глазных щупалец. Эффект сохранялся в течение многих дней. Однако он проявлялся только в той обстановке, в которой применялось болевое раздражение (на шаре в воде), и отсутствовал, когда улитка тестировалась в других условиях — на стеклянной крышке аквариума. Применение нейротоксина, избирательно нарушающего работу серотонинергических нейронов, устраняло его проявление. Это одна из форм долговременной сенситизации, проявляющая себя как обстановочный условный рефлекс и требующая участия модулирующих серотонинергических нейронов. Предполагается, что данный обстановочный условный рефлекс реализуется на входах модулирующих нейронов, которые являются нейронами «состояния» [Балабан и др., 1992].
Модулирующие нейроны участвуют в процессе научения (см. гл. 15), изменяя проводимость синапса на пресинаптическом уровне. Эффект пресинаптической пластичности, создаваемый модулирующими нейронами, может быть начальным звеном в цепи событий, приводящих к пластической модификации поведения, связанной с выработкой условных рефлексов. Безусловное подкрепление как очень сильный раздражитель нейрона также возбуждает модулирующие нейроны и тем самым инициирует процесс пресинаптических изменений. За последним следует этап клеточных и молекулярных преобразований постсинаптического нейрона, характеризующий ассоциативное обучение.
Однако пресинаптическое облегчение, создаваемое модулирующими нейронами, может возникать и вне ситуации ассоциативного обучения только за счет активации модулирующих нейронов сильными сенсорными раздражителями. Э. Кэндел и Р. Хокинс [1992] рассматривают его как результат «премодулирующего совпадения», которое на пресинаптическом уровне повышает проводимость, а само это повышение зависит от активности. Другое название этого явления — сенситизация, или неассоциативное обучение. Оно отличается от ассоциативного обучения, основанного на «препостсинаптическом совпадении» (рис. 8.1).
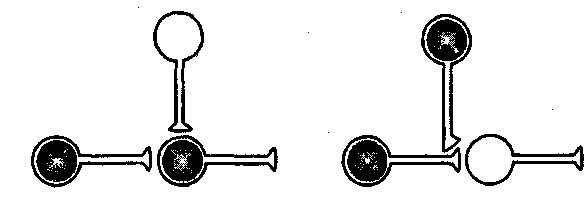
Рис. 8.1. Два механизма изменения силы связи между нейронами. Один требует совпадения активности пресинаптического и постсинаптического нейронов во времени. Другой основан на совпадении активности пресинаптической клетки и модулирующей. Нейроны, участвующие в научении, заштрихованы [Кэндел, Хокинс, 1992]
Эффект пресинаптического облегчения с участием модулирующих нейронов хорошо изучен при формировании оборонительного рефлекса у виноградной улитки. Внутриклеточная стимуляция ее модулирующих нейронов, которые представлены пулом серотонинергических клеток педального ганглия, вызывает в командном нейроне оборонительной реакции появление деполяризационной волны и увеличение амплитуды суммарного ВПСП. Эти изменения превращают сенсорное подпороговое раздражение командного нейрона в пороговое, что выражается в появлении у него спайковой активности.
Описано два механизма длительного пресинаптического облегчения (фасилитации) под влиянием серотонина за счет фосфорилирования белков двух типов ионных каналов: кальциевых и калиевых (рис. 8.2).
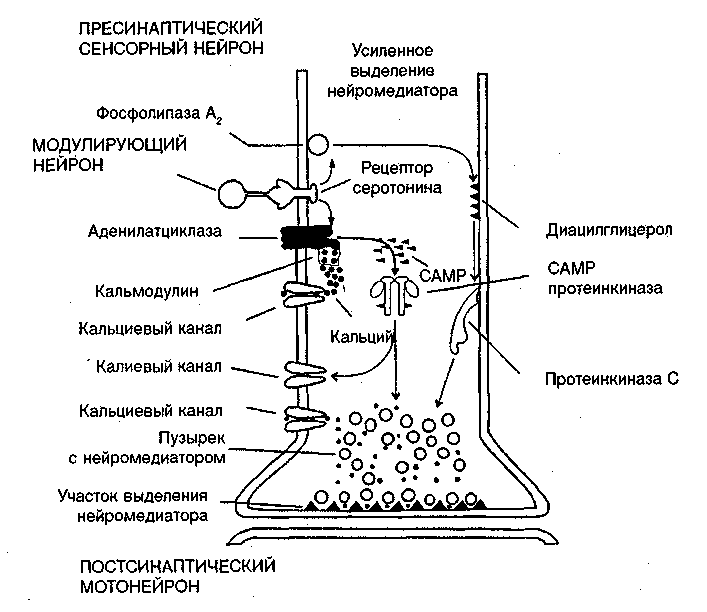
Рис. 8.2. Схема молекулярных реакций в пресинаптическом нейроне под влиянием возбуждения модулирующего серотонинергического нейрона — научение, зависимое от активности [Кэндел, Хокинс, 1992]
Участие кальциевых каналов в механизме повышения проводимости через синапс выглядит следующим образом. Во время потенциала действия ионы кальция и натрия входят в клетку, а ионы калия выходят из нее. Когда нейрон находится в активном состоянии, в нем увеличивается уровень содержания ионов кальция. Кальций, вошедший в клетку, действует на нее после того, как связывается с белком — кальмодулином. Этот комплекс так же, как и серотонин, активирует аденилатциклазу. За счет двойного воздействия на нее резко увеличивается образование CAMP из аденозинтрифосфата (АТФ), который служит в клетке универсальным источником энергии для биохимических процессов; CAMP действует как вторичный посредник и активирует протеинкиназу. Протеинкиназа фосфорилирует рецепторные белки кальциевых каналов (присоединяет к ним фосфор). Это переводит кальциевые каналы в активное состояние — в клетку входит больше кальция, что увеличивает выделение из нее медиатора. Одновременно протеинкиназа фосфорилирует белки калиевых каналов. Это ведет к уменьшению проводимости для ионов калия, т. е. к уменьшению калиевого тока, что ограничивает калиевую гиперполяризацию, следующую за ВПСП и потенциалом действия. Таким образом, потенциал действия продлевается, а это, в свою очередь, увеличивает продолжительность активного состояния кальциевых каналов.
Особую функцию в модуляции возбудимости нервной системы выполняют пейсмекерные нейроны (см. гл. 1). Будучи активированными, они своими длительными разрядами могут повышать проводимость сигнала через синапс. Специальный тип пейсмекерных нейронов, генерирующих разряды на частоте гамма-колебаний, которые, по-видимому, отражают колебания кальциевого тока на дендритах клеток таламуса, синхронизирует активность больших массивов нейронов, объединенных общими резонансными свойствами.
Эмоциональный стресс.
Под эмоциональным стрессом в самой общей форме понимают общую системную реакцию организма, развивающуюся на действие стресс-факторов (стрессоров). Оценка какого-либо воздействия, ситуации как негативных (отвергаемых) при невозможности избавления от них или неподготовленности механизмов "защиты" позволяет рассматривать эти факторы в качестве стрессоров. При этом надо иметь в виду, что не само по себе воздействие, а именно отношение к нему, негативная его оценка с сенсорной, психологической или социальной точки зрения является причиной эмоционально-стрессовой реакции организма (Вальдман и др., 1979). Главная сущность стресс-сигнала в его информативном значении, поэтому он оценивается в категории субъективного, личностного к нему отношения. Следовательно, типологические свойства субъекта, особенности эмоционального восприятия и эмоциональных свойств личности и определяют тип реагирования и направленность поведения. В развитии стресса различают три стадии: 1. Стадия тревоги, возникающая при внезапном действии стрессора, проходит в форме шока как результата сильнейшего эмоционального возбуждения. 2. Стадия резистентноcти, подготавливающая организм к повышенной устойчивости к действию вредных факторов. Адаптация в основном осуществляется системой гипоталамус - кора надпочечников, так называемыми адаптивными гормонами, и прежде всего аденокортикотропным гормоном. 3. Стадия истощения, возникающая при продолжающемся воздействии стрессоров. Эта стадия еще именуется состоянием психической дезадаптации. Приведем следующий пример из экспериментов с животными (М.А. Батуева, 1985). В одном ограниченном жилом помещении находятся два однопометных кролика, один из которых характеризуется преобладанием пассивно-оборонительных реакций, а другой - реакциями активного нападения. Иными словами, имеет место типичный случай эмоционально-психологической несовместимости. Изоляция животных друг от друга позволила у каждого животного выработать двитательнопищевые рефлексы. По критерию стабильности рефлексов и их количественных параметров животные не отличались друг от друга. В один из опытных дней животные были сведены вновь в общее жилое помещение. Спустя три дня совместного проживания у одного из кроликов (пассивного) исчезли условные рефлексы, тогда как у другого (активного) все осталось по-прежнему. Еще через три дня животные были вновь разведены в разные жилые клетки. Ежедневное тестирование показало постепенное в течение 6 дней медленное восстановление условных рефлексов у пассивного кролика. В данном случае в качестве стрессора выступает особь того же вида при наличии антагонистических взаимоотношений. Максимальное напряжение механизмов защиты возникает в системе при прагматической неопределенности, угрожающей ситуации, при невозможности осуществления готовых программ деятельности, при необходимости срочного подбора новых вариантов поведенческих программ. Наиболее реактивной из структур мозга является кора больших полушарий, а в ней - митохондрии и синапсы, особенно тормозные, а в самой нервной клетке - ферментные системы энергетического и медиаторного обмена. Психотропные вещества, вмешиваясь в процессы "химической интеграции" нейронов, могут менять информационное значение сигнала для данного индивида. Длительное и эффективное психофармакологическое лечение изменяет и тот памятный след, который является основой психотравмирующей ситуации, и ведет к полному или частичному восстановлению нормальных психических функций. Одним из эффективных терапевтических приемов является активизация собственных компенсаторных механизмов мозга, направленных на устранение или ограничение патогенных процессов (М.М. Хананашвили, 1972).